2. fejezet - Az emberi látással kapcsolatos alapismeretek
Szemünk a legfontosabb érzékszervünk. Agyunk a külvilágból származó információk 90%-át a színes látás útján szerzi, és ez az érzékünk alakítja leginkább külvilágról alkotott képünket. Látószervünk hétköznapi életünkben betöltött fontos szerepe tehát vitathatatlan. Mindezeken túl ahhoz, hogy a fényt, mint az elektromágneses sugárzás számunkra érzékelhető tartományát (2.1. ábra) műszaki szempontból is kezelni tudjuk, elengedhetetlenül szükséges, hogy megismerkedjünk a színes látás rendszerének alapvető elemeivel, jellemzőivel és működési módjával.
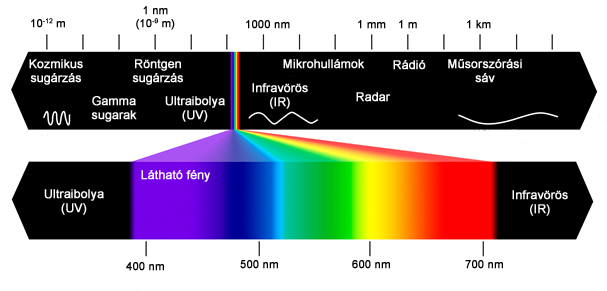
Ennek első lépése a fény, mint fogalom definiálása. Attól függően, hogy a látórendszerünk működésének mely szintjén vizsgálódunk, három különböző definíció is megadható.
Eszerint a fény (az MSZ 9620-as fénytechnikai terminológiának megfelelően)
-
Fizikai szempontból:
-
Meghatározott hullámhosszú sugárzás, inger
-
-
Fiziológiai szempontból:
-
A látás érzékszervében a fényinger által kiváltott érzet
-
-
Pszichológiai szempontból:
-
Az agykérgi látóközpontban létrejött észlelet
-
A fényjelek, jelzések és kijelzők az autóiparban számos területen bírnak igen nagy fontossággal. Számtalan fénytechnikai megoldás található a gépjárművek belterében, amelyek éjszakai vezetés közben is ellátják a vezetőt és az utasokat a szükséges információkkal. Még talán ennél is fontosabb a fénytechnika fényszórókban és a helyzet- vagy szándékjelző berendezésekben betöltött szerepe, hiszen e funkciók elsődleges feladata a balesetek megelőzése, és a biztonságos közlekedés lehetővé tétele.
Ahhoz, hogy érdemben beszélhessünk a fényről, valamint alkalmazásáról az autóipar egyes részterületein, meg kell határoznunk, hogy az érzékelés mely szintjén zajlanak a technikai szempontból fontos folyamatok. Eszerint jelen jegyzet leginkább érzet formájában fog foglalkozni a fénnyel, esetenként röviden érintve a fizikai és a pszichológiai definíció által körülhatárolt területeket. Ebben a fejezetben sorra fogjuk venni az emberi szem felépítésével, a színes látás működésével és a színek kvantitatív jellemzésével kapcsolatos legfontosabb ismereteket, valamint két, vezetéstechnikai szempontból kritikus részterületet, a mezopos látás sajátosságait és a káprázás, valamint vakítás kérdéskörét.
2.1. Az emberi szem
Szemünk rendkívül összetett optikai rendszer. Fontosságát jól mutatja, hogy az egyedfejlődés során eme szervünk éri el leghamarabb a kifejlett állapotot. Egy ötéves gyermek szeme már teljesen kifejlett, mérete és fizikai felépítése nem változik tovább. Az is figyelemre méltó, - és jól mutatja a konstrukció kifinomultságát - hogy az evolúció során egymástól független módokon, de sokszor nagyon hasonló felépítéssel több esetben is alakult ki látószerv. Nem véletlen tehát, hogy a Földünkön élő állatfajok jelentős többsége rendelkezik fényingerek feldolgozására alkalmas szervvel.
Az emberi szem – akárcsak a legtöbb összetett látószerv - felépítését tekintve két részre bontható. Optikai rendszerre, amely a fényingerek összegyűjtéséért és fókuszálásáért felelős, valamint neurális rendszerre, amely az optikai elemeken keresztüljutó ingerek előfeldolgozását, és az agyba való továbbítását végzi.
2.1.1. A szem optikai rendszere
Ha a fény terjedési iránya által meghatározott sorrendben vizsgáljuk szemünk részeit, az első optikai elem a szaruhártya (2.2. ábra/1) vagy más néven cornea. Optikai szempontból ez, a szemgolyónk elülső felén található kidomborodó terület rendelkezik a legnagyobb törőerővel, ezáltal ennek alakja határozza meg leginkább látásunk minőségét. Az előző fejezetben taglalt törési törvény ismeretében nem meglepő, hogy optikai szempontból ez a legnagyobb hatással bíró elem . Mivel a szemlencse által képzett optikai határfelület egyik oldalán levegő, a másik oldalán pedig a szaruhártya anyaga található, könnyedén belátható, hogy a törésmutatók közötti különbség itt lesz a legnagyobb az egész rendszeren belül. Ez az oka annak is, hogy lézeres szemműtétek során a szaruhártya görbületének megváltoztatásával érnek el látásjavulást.
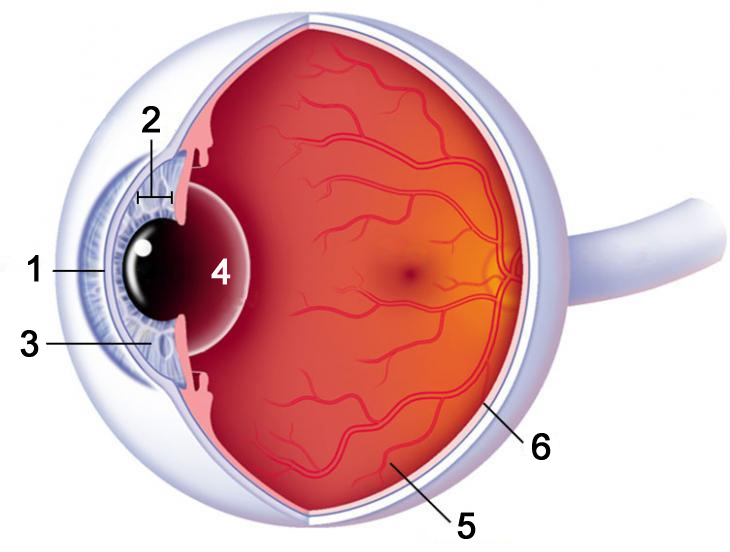
A szaruhártya mögött az elülső csarnok található (2.2. ábra/2), amelyet vízszerű, viszkózus folyadék – csarnokvíz - tölt ki. Ennek szerepe a szaruhártya alakjának megtartása, illetve a szemlencse és a szaruhártya tápanyagokkal való ellátása, hiszen ezeket nem hálózzák be erek. Ennek megfelelően a csarnokvíz folyamatosan cserélődik, a csarnokzugban található nyíláson keresztül elfolyik, és újratermelődik. Ha a kivezető nyílás valamilyen okból eltömődik, a csarnokvíz nyomása megemelkedik, amely maradandó látáskárosodáshoz vezethet – ez a zöldhályog.
Az elülső csarnokban még a szemlencse előtt található a pupilla, vagy más néven íriszhártya (2.2. ábra/3), amelynek elsődleges feladata a szem további részeibe bejutó fény mennyiségének szabályozása. Ezt adaptációs mechanizmusnak nevezzük, kiváltó oka pedig a környezetben beálló fénysűrűség változás. A pupilla tágulását és szűkülését a szemben található simaizmok teszik lehetővé, a folyamat szabályozása pedig a retinán található, – többek közt – erre a feladatra specializálódott neuronok által gyors magnocelluláris idegpályákon közvetített jelek révén történik. A hártya sejtjei pigment anyagot is tartalmaznak. Ezen anyagoknak köszönhetően létezhetnek különböző színű szemek, fiziológiai szerepük azonban jelenlegi ismereteink szerint nincs. A környezet fénysűrűségéhez való adaptációval a szem optikai rendszerének tulajdonságai is megváltoznak, hiszen a pupillaátmérő változása egyfajta rekeszelésnek tekinthető.
A pupilla mögött található a szemlencse (2.2. ábra/4), amelynek legfontosabb feladata látószervünk akkomodációs képességének biztosítása. Az akkomodáció során szemünk optikai rendszere alkalmazkodik a vizsgált objektum és nézőpontunk közötti távolsághoz. Ez a szemlencse alakjának módosításával érhető el. Ha a szemlencse görbületi sugara csökken, nézőpontunkhoz közelebbi tárgyakra fókuszálunk, ha a görbületi sugár nő, a távolabbi tárgyak képe vetül élesen a retinánkra. A görbületi sugár módosítását a szemlencse hagymához hasonló héjas szerkezete, valamint a hozzá tapadó ciliaris izomzat, az úgynevezett sugártest teszik lehetővé. Az akkomodációs képesség az életkor előrehaladtával csökken, valamint egyénenként is változó lehet.
A fentieken túl szemlencsénk egyfajta szűrőként is funkcionál, megvédve retinánkat a káros, nagy energiájú UV sugárzástól. Ehhez a lencse anyaga a 400 nm-nél rövidebb hullámhosszú sugárzás jó részét elnyeli, vagyis a látható tartomány alsó határát tulajdonképpen a szemlencse transzmissziós karakterisztikája határozza meg. Az elnyelt UV sugárzás hatására a lencse anyaga életünk során fokozatosan bebarnul – sötétedik –, ezzel egyfajta „beépített napszemüvegként” működik. Ezzel párhuzamosan csökken a szembe jutó kék fény mennyisége, amely jelenséget agyunk a neurális feldolgozási folyamatok révén részben korrigál, így a lassú, de folyamatos változás észrevétlen marad. Mindazonáltal a jelenség alapvető különbségeget okoz a fiatal és az idősebb emberek fényérzékelésében, amelyet a tervezési folyamatok során szem előtt kell tartani. A szemlencse – jellemzően idős korban – akár teljesen átlátszatlanná is válhat, ezt nevezzük szürkehályognak. A szürkehályog ma már rutinszerűnek tekinthető műtéti eljárással gyógyítható, amelynek során a páciens szemlencséjét eltávolítják, és egy jellemzően polimer anyagú műszemlencsével helyettesítik. A műszemlencse nem tesz lehetővé akkomodációt, de mivel idős korra ez a képesség már úgyis csak csekély mértékben funkciónál, a páciensek számára ez nem jár különös hátrányokkal.
Szemünk utolsó, a leképzésben szereppel bíró eleme a hátulsó csarnokban található üvegtest (2.2. ábra/5). Ezt a hátulsó csarnokvíz tölti ki, amely átlátszó, kocsonyás anyag. Ez adja szemünk szerkezeti stabilitását, és az optikai szempontból előnyös gömbformát. A hátulsó csarnokvíz nem cserélődik, így törmelékek halmozódnak fel benne az idő előrehaladtával, és apró – néha zavaró – foltokként jelennek meg látóterünkben.
A szemgolyónk hátulsó falát majdnem teljes egészében beborító idegsejtek alkotják a retinát, más néven recehártyát (2.2. ábra/6). Ennek megfelelően a retina sokkal inkább tekinthető agyunk, mintsem szemünk részének. Felépítésével és működésével később külön alfejezetben részletesen foglalkozunk.
2.1.2. A szem leképzési hibái
Mint a legtöbb optikai rendszer a szem is rendelkezik bizonyos leképzési hibákkal, aberrációkkal. Ezek lehetnek a rendszer felépítéséből következő hibák vagy olyan elváltozások, amelyek egyénspecifikusak, esetleg az öregedés folyamatával jelennek meg. Ezek közül néhányat már a korábbiakban említettünk, akad azonban olyan is, amely behatóbb leírást igényel.
Az emberi szem legjellemzőbb leképzési hibája az ametropia, vagyis a retinára való fókuszálás hibája. Ez azt jelenti, hogy a szem optikai rendszere nem éles képet vetít a retinára, rontva ezzel a látórendszer felbontóképességét. Az ametropia a legtöbb embert érinti, hiszen a populáció csak nagyon csekély része rendelkezik tökéletes látással - emmetropiával. Ez a leképzési hiba több okra is visszavezethető, melyek közül a legjellemzőbb, hogy a szem optikai elemeinek alakja eltér az ideális felületi formától. Az ametropia két jellemző típusa a miopia (2.3. ábra/1) vagy közellátás, illetve a hiperopia (2.3. ábra/2) vagy távollátás.
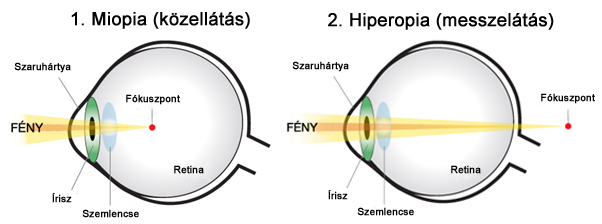
Távollátásról akkor beszélünk, ha a szaruhártya és a szemlencse törőereje együttesen nem elég nagy ahhoz, hogy a környezet képe megfelelően fókuszálódjon a retinára. Ilyenkor éles kép - korrekció nélkül - a retina mögött keletkezik, ami leginkább a közelebb eső tárgyak szemlélése esetén okoz problémát. A hiperopia korrekciója a szem elé helyezett, megfelelő törőerejű sugarú gyűjtőlencsével történik.
A miopia, avagy közellátás a hiperopia ellentéte. Ilyenkor a szemlencse és a szaruhártya együttes törőereje túl nagy, az éles kép a retina előtt keletkezik, így a távolabb lévő tárgyak éles leképzése válik problémássá. Korrekciójához megfelelő törőerejű szórólencse szükséges.
Az akkomodációs képesség időskori csökkenése szintén ametropiához vezet, hiszen a szemlencse rugalmasságának csökkenésével beszűkül az a távolságtartomány, amelyen belül a látórendszer éles képet tud a retinára vetíteni. Az ilyen típusú szemhiba elnevezése a presbiopia.
A fentieken túl a látásélességre ható leképzési hiba a szférikus aberráció. Ez abból adódik, hogy a tökéletes látással rendelkező egyének szemében található optikai elemek felületei sem alkalmasak a tökéletes képalkotásra, csak közelítik azon negyedrendű felületek alakját, amelyek a teljes képtartományon tökéletes leképzést biztosítanának. Gyakorlati szempontból ez a hiba azonban kevésbé fontos, mert a retina felépítésében tapasztalható sajátosságok jelentősen csökkentik e hibának negatív hatásait. Erről a későbbiekben bővebben is szót ejtünk.
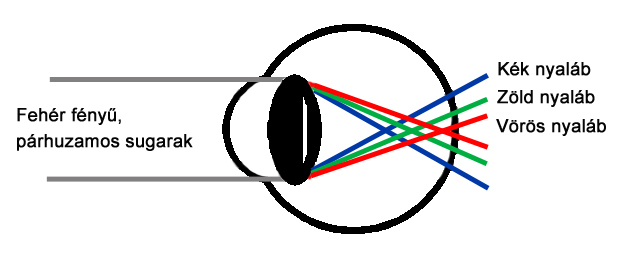
A látásélességet befolyásoló, fent említett aberrációs jelenségeken kívül megemlítendő még a szem optikai rendszerének kromatikus aberrációja is. A kromatikus aberráció oka, hogy az egyes optikai elemek törőereje nem független a beeső fény hullámhosszától (2.4. ábra), ennek következtében a különböző színekre különböző fókusztávolsággal bír az optikai rendszer. Ennek azért van jelentősége, mert heterokromatikus fényingerek esetén, főleg ha az inger kvázi-monokromatikus sugárzások additív keverékéből áll össze - például az autók műszerfalán alkalmazott kijelzők esetében - Az egyik színre akkomodálódott látórendszer nem képes a jellemzően más színű ingerek finom részleteinek megkülönböztetésére, hiszen azok képe életlenül vetül a retinára.
2.1.3. A retina felépítése és működése
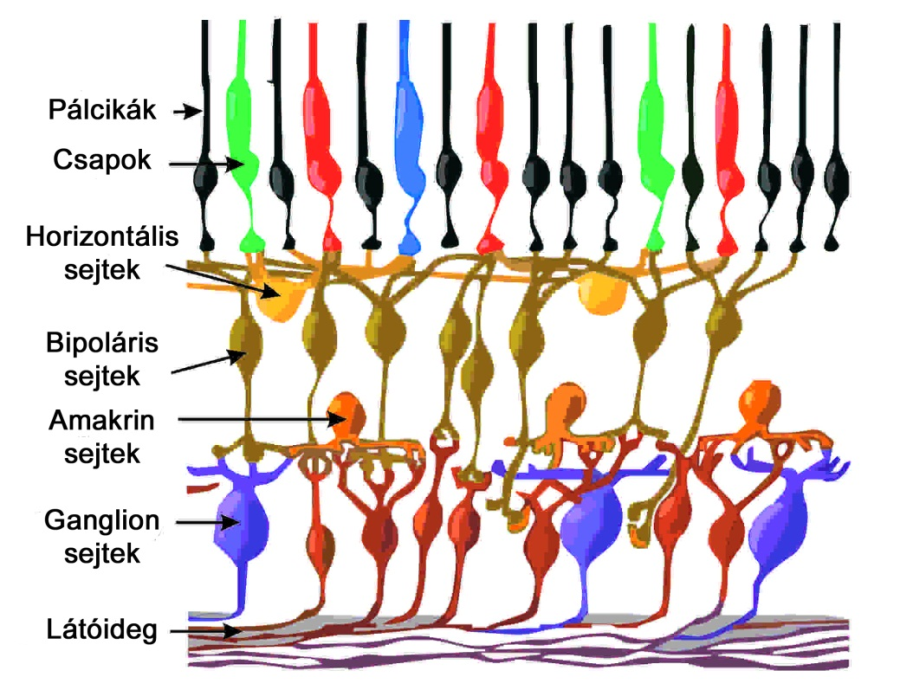
A retinát több, egymással összeköttetésben álló sejtsor alkotja (2.5. ábra). Az ingerek terjedésének sorrendjében haladva megtaláljuk a fényérzékeny receptorok, azaz fotoreceptorok rétegét, majd a horizontális sejtek sora következik. Ezt a bipoláris és amakrin sejtek rétege követi, legvégül pedig a ganglion sejtek csoportjai következnek. Jelen alfejezetben sorra vesszük a retina fontosabb sejtjeit, azok működését, továbbá a látásérzékelés retinális feldolgozási fázisának fő elemeit. Fontos megjegyezni, hogy a retinát alkotó sejtek némelyikének teljes funkcionalitása a mai napig nem ismert, és habár a látás és színlátás neurális folyamatairól már nagyon komplex modellekkel rendelkezünk, még bőven akad kutatni és felfedezni való ezen a területen. Az mindenesetre biztos, hogy a retina látásérzékelésünk első bástyája, a frontvonalban pedig a fényérzékeny csapok és pálcikák helyezkedek el.
A fotoreceptorok az úgynevezett transzdukciós mechanizmus során alakítják a vizuális ingereket agyunk számára feldolgozható idegi jelekké. A receptorsejtek felépítése bipoláris (2.6. ábra), külső és belső szegmensre bonthatóak. A belső szegmensben helyezkedik el a minden idegsejtre jellemző szinaptikus végződés, amelynek feladata a többi idegsejttel való kapcsolat kialakítása. Szintén a belső szegmensben találhatóak a hagyományos citoplazmikus sejtszervecskék, többek közt az örökítő anyagot tartalmazó sejtmag és a sejt energiaellátásáért felelős mitokondriumok, valamint a transzdukciós mechanizmusban szerepet játszó nátriumion pumpák. A külső szegmensben találhatóak a fényérzékeny opszin molekulák.
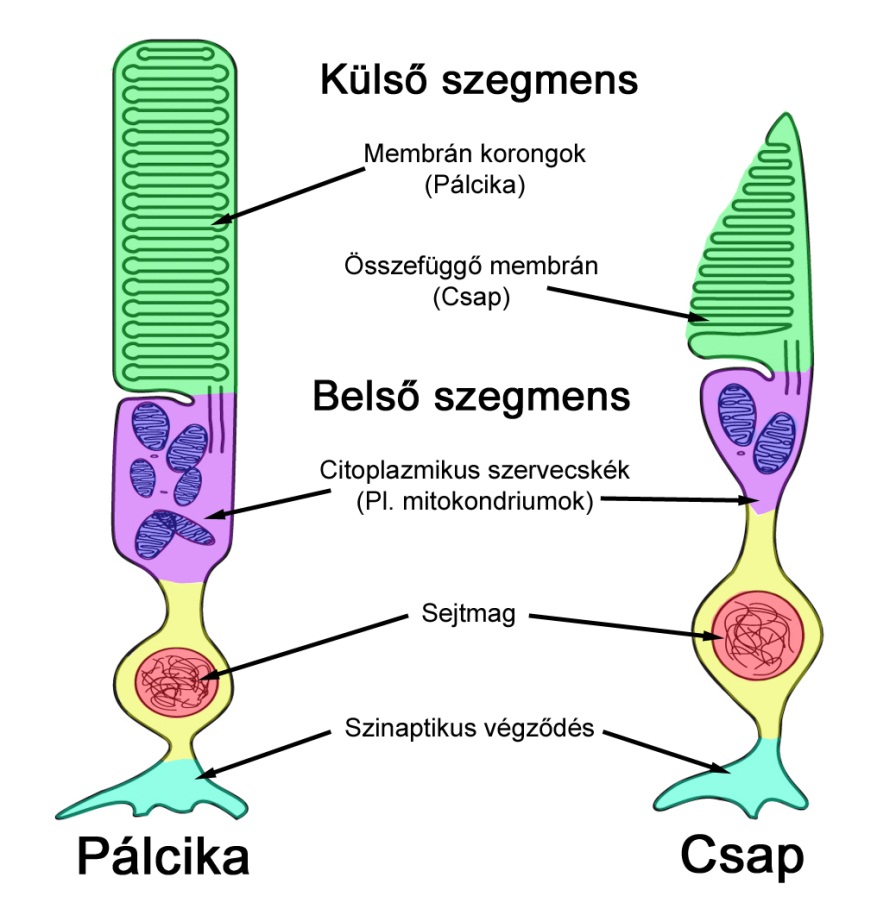
A külső szegmens szerkezete speciális, membrános kialakítású. A pálcikák esetében zárt, korong-szerű membránok alkotják, a csapok esetében pedig harmonikaszerűen egymásra rétegződő, összekapcsolódó membránszalag alkotja ezt a sejtrészt. A membrános felépítés mindkét receptortípusnál nagymértékben megnöveli a sejtfal fajlagos felületét a külső szegmensben, amelynek nagy szerepe van a folyamat működése szempontjából. A membránokhoz kapcsolódó opszin molekulák a beérkező fény hatására izomerizálódnak, ezzel inkompatibilissé válva a membránok csatlakozási pontjaihoz. Az izomerizálódott opszinok leválnak a membránokról, és elindítják a fototranszdukciós kaszkádnak nevezett folyamatot, amelynek során nátriumion csatornák záródnak be a sejtfalon. Ennek hatására a sejt belseje és környezete között potenciálkülönbség alakul ki, amelynek következtében a sejt külső felületén elektromos jel keletkezik. Mivel a keletkező áram arányos a bezáródó ioncsatornák mennyiségével, amely pedig a beérkező fény hatására izomerizálódó molekulák mennyiségének függvénye, a fotoreceptorok által közvetített jel minden pillanatban arányos lesz a beérkező fotonok mennyiségével. Ez jelentősen eltérővé teszi ezen sejteket a többi, jellemzően akciós potenciállal rendelkező idegsejttől, amelyek a jeltüskék sűrűségében kódolva, kvázi-digitális módon továbbítják és dolgozzák fel a jeleket. A retinán a jelfeldolgozás és továbbítás csak a látókéreg előtti utolsó „állomáson”, a ganglion sejtek kimenetén lesz frekvenciamodulált.
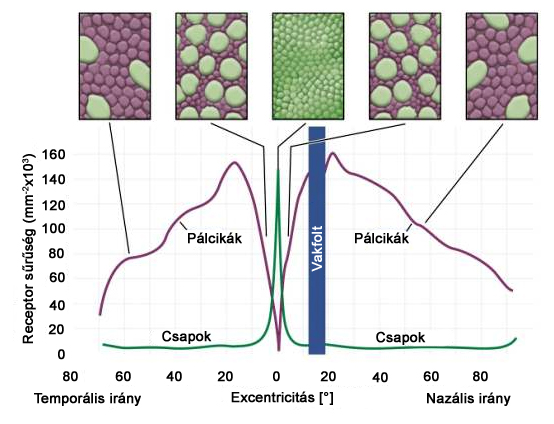
A fotoreceptorok közül a pálcikák az éjszakai, vagy kis megvilágítású környezetben működő, úgynevezett scotopikus látásért felelősek. Egy típusuk van, érzékenységi maximumuk nagyjából az 505 nm-es hullámhossz környékén található. Számuk hozzávetőlegesen 106 millió. A csapok és pálcikák eloszlását a retinán a 2.7. ábra mutatja. Ezen jól látszik, hogy a pálcikák jellemzően a retina perifériális részén helyezkednek el. Ennek oka, hogy mivel az ember jellemzően a nappali órákban aktív, éjszaka jórészt csak veszélyérzékelésre használjuk látásunk. Ehhez a perifériális látóterünkben tapasztalható változások fontosabbak, így itt a jeleket sokkal gyorsabb idegpályákon továbbító pálcikák túlsúlya indokolt.
Nem csak a gyors jeltovábbítás teszi azonban alkalmassá a pálcikákat a kis megvilágítású környezetben való működéshez, hanem a csapokhoz képest jóval nagyobb érzékenységük. A pálcikák külső szegmensében található opszin molekula a rodopszin, ez egy meglehetősen instabil vegyület, izomerizációja nagyobb valószínűséggel megy végbe - azonos beérkező foton mennyiségnél -, mint a csapok opszinjai esetében. Ennek következtében a pálcikákkal jóval alacsonyabb intenzitású fényingereket is képesek vagyunk érzékelni. A csapok és pálcikák retinán vett eloszlásának, és az érzékenységükben tapasztalható különbségek szemléltetésére a legjobb példa a halvány csillagok megfigyelése. Ha egy tiszta éjszakán felpillantunk az égre, sok olyan csillagot találunk, amelyek nem láthatóak, ha közvetlenül rájuk tekintünk, kissé „mellénézve” azonban érzékelhetővé válnak. Ennek a jelenségnek a magyarázata, hogy látómezőnk közepén a színes látásért felelős csapok helyezkednek el, amelyek kevésbé érzékenyek, mint a pálcikák, így ezek a halovány csillagok érzékelésére nem képesek. Ha a perifériális látómezőnkbe pozícionáljuk a vizsgálni kívánt objektumot, annak fénye jó eséllyel a pálcikákra vetül, amelyek már kellőképpen reszponzívak ahhoz, hogy ezt a csekély fényingert is érzékelhetővé tegyék agyunk számára.
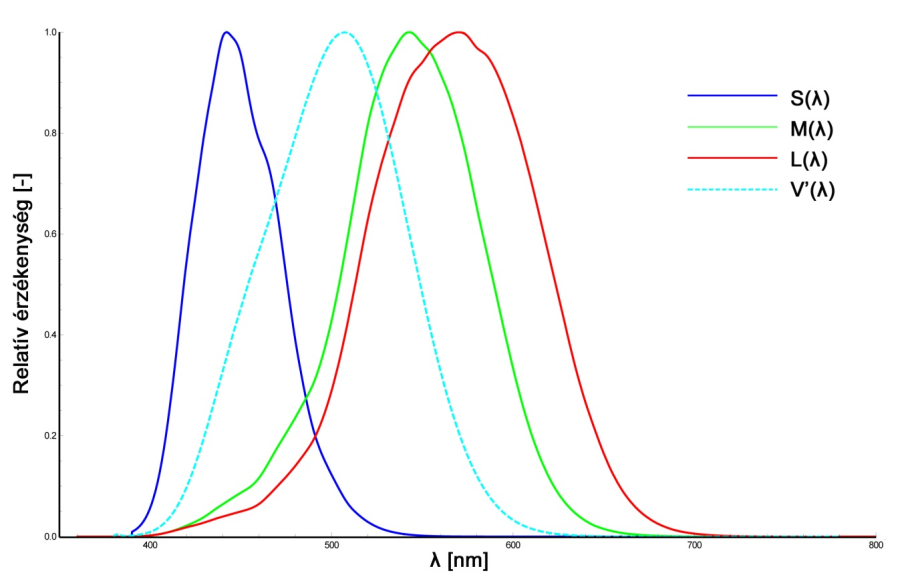
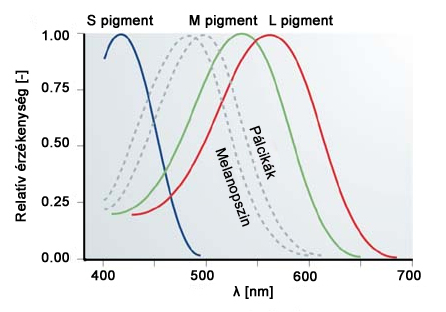
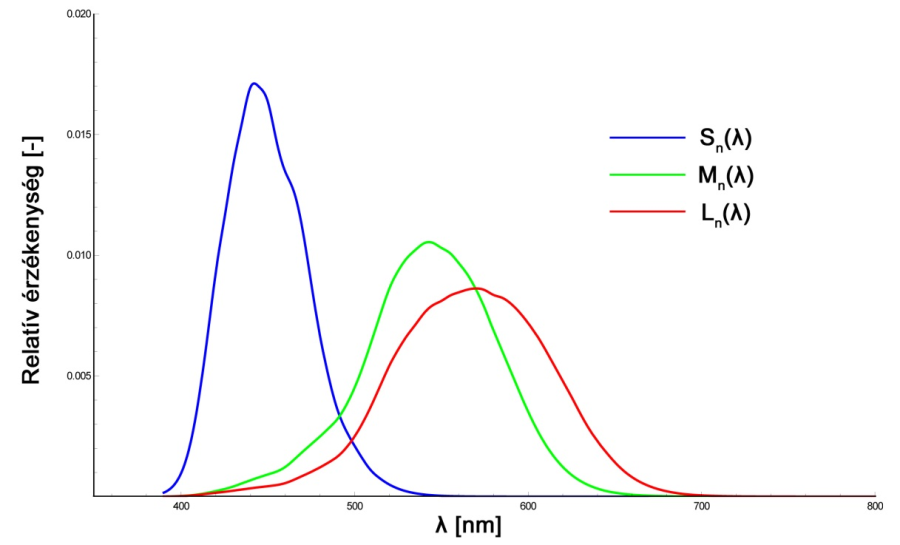
A csapok a nappali, vagy fotopikus látásért felelős fotoreceptorok. Kevésbé érzékenyek, mint a pálcikák, hiszen nappali körülmények között jóval nagyobb intenzitású fényingerek érnek bennünket. A csapoknak három típusa van, ami a gyakorlatban azt jelenti, hogy az egyébként azonos felépítésű csapsejtek kissé eltérő opszin molekulákat termelnek. Eszerint megkülönböztetünk kék-érzékeny, más néven tritos, zöld-érzékeny vagy deuteros, illetve vörös-érzékeny, azaz protos elnevezésű csapokat. Ezen receptorok jeleinek összevetésével válik lehetségessé a színek érzékelése, ennek megfelelően a csapsejtek, illetve a bennük termelődő és fény hatására lebomló fotoszenzitív molekulák alkotják színlátásunk fiziológiai alapjait. A tritos, deuteros és protos receptorok érzékenységi maximumai rendre 420-440 nm, 534-545 nm valamint 564-580 nm közé esnek. A csap receptorok (S(λ), M(λ), L(λ)), valamint a pálcikák (V’(λ)) spektrális érzékenységi karakterisztikáját a 2.8. ábra mutatja be.
Ahogy azt a 2.7. ábra is mutatja, a rendelkezésünkre álló hozzávetőlegesen 6 millió csap jellemzően a retina közepén, a foveolának vagy látógödörnek is nevezett gödröcskében található meg nagy mennyiségben, a retina perifériális részein számuk meglehetősen alacsony. A foveola a retinának a szem optikai tengelyéhez legközelebb eső része. Látórendszerünk optikai tulajdonságai ezen a területen a legjobbak, az optikai rendszer szférikus aberrációjának - vagyis az optikai tengelytől távol eső sugarak leképzési hibájának - hatása itt a legkisebb a látásélességre. Ez az elrendezés lehetővé teszi, hogy a szférikus aberrációból adódó leképzési hibák ne terheljék nagymértékben a látórendszer felbontóképességét.
Megjegyzendő, hogy a retina felől az agyba távozó idegrostok száma a látóidegben mintegy két nagyságrenddel kevesebb, mint a retinán megtalálható fotoreceptorok együttes száma. Ennek oka az, hogy a retinán található egyéb sejtek – amakrin, bipoláris és ganglion sejtek – feldolgozzák a receptorokból származó ingerületeket, és az agyba már nem a receptorokból származó közvetlen primer jelek jutnak. Valószínű, hogy a foveola területén elhelyezkedő nagyszámú csap csaknem mindegyike rendelkezik direkt az agyba vezető idegpályával, azonban a perifériális részeken a receptorok jelei nagy számban összegződnek.
A bipoláris, horizontális és amakrin sejtek az úgynevezett interneuronok. Feladatuk az ingerületek továbbítása a fotoreceptoroktól a ganglion sejtek felé, valamint a fotoreceptorokat összefűző receptormezők kialakítása, és megfelelő bemenetek képzése a ganglion sejtek számára. Ganglion sejt és fotoreceptor között két útvonal lehetséges:
-
Direkt (vagy vertikális): Fotoreceptor – Bipoláris sejt – Ganglion sejt
-
Indirekt (vagy horizontális): Fotoreceptor – / Horizontális sejt / – Bipoláris sejt – / Amakrin sejt / – Ganglion sejt
Ahol:
|
„-„ : Rétegek közti direkt kapcsolat |
||
|
„/” : Laterális, indirekt kapcsolat |
Indirekt útvonalon a jelfolyamba bekapcsolódnak a horizontális és az amakrin sejtek is. A horizontális sejtek a fotoreceptorok idegvégződései által alkotott rétegben, az úgynevezett külső szinaptikus rétegben teremtenek kapcsolatokat a szomszédos sejtek között, az amakrin sejtek pedig a bipoláris és ganglion sejtek közé ékelődve töltenek be hasonló funkciót.
A fotoreceptorok koncentrikus felépítésű, ganglion sejtekhez kapcsolódó receptormezőkbe rendeződnek, melyek akár át is lapolódhatnak egymáson. A pálcikák nagyméretű, homogén mezőket alkotnak, közvetlen kapcsolatban pedig csak egyféle bipoláris sejttel állnak. Egy-egy pálcikákat összekapcsoló bipoláris sejthez hozzávetőlegesen 15-30 receptor tartozik. Ezek a bipoláris sejtek soha nem állnak közvetlen szinaptikus összeköttetésben ganglion sejtekkel, a jelfolyamba minden esetben amakrin sejtek ékelődnek. Hasonlóan a pálcikák és a hozzájuk kapcsolódó bipoláris sejtek kötegelődéséhez, egy-egy amakrin sejthez is több – nagyjából sejtenként 20 – pálcikákat összekapcsoló bipoláris kapcsolódik. A konvergencia a ganglion és amakrin sejtek között még ennél is nagyobb arányú lehet, esetenként egy-egy ganglion sejthez több mint száz pálcika jeleket továbbító amakrin sejt is tartozhat. Mindezekből könnyen kiszámítható, hogy a pálcikákra legjellemzőbb útvonalat követve egyeten ganglion sejthez akár sok tízezer receptor jele is befuthat. Ez a nagymértékú, retinális rétegeken átívelő transzverzális konvergencia komoly szerepet játszik a pálcikák dominálta szkotópikus, azaz éjszakai látás nagymértékű érzékenységében.
A csapok alkotta receptormezők felépítése nem homogén, centrális és perifériális részből áll (2.9. ábra). A centrális és perifériális szegmensek közötti eltéréseket a kétféle, on és off típusú bipoláris sejtek alakítják ki. Ennek megfelelően a bipoláris sejtek úgy is felfoghatók, mint egyféle előjelképző állomások a ganglion sejtek és csapok, vagy csapok csoportjai között.
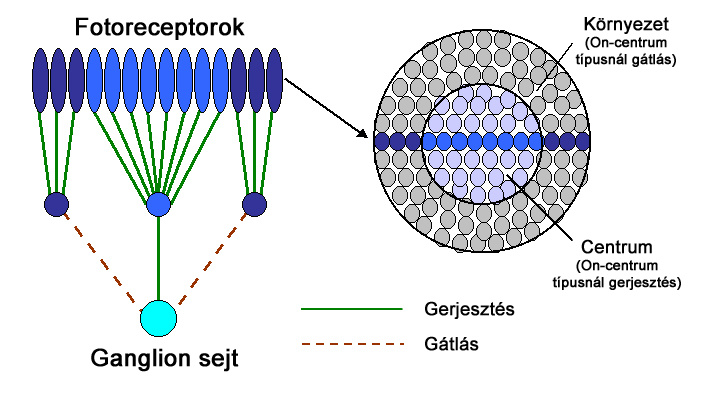
Attól függően beszélhetünk on-centrum és off-centrum receptor mezőről, hogy a receptor mezők centrumai milyen típusú bipoláris sejten keresztül kapcsolódnak a hozzájuk tartozó ganglion sejthez(2.10. ábra). Ennek megfelelően a ganglion sejtek viselkedése az előttük lévő bipoláris sejtek viselkedését tükrözik, de egyes esetekben horizontális és amakrin sejtek is módosíthatnak a jelfolyamon. On-centrum esetben a receptor mező perifériális részének ingerlése gátolja, centrális része pedig tüzelésre – jelkibocsájtásra - készteti a ganglion sejtet. Off-centrum esetben a hatásmechanizmus éppen ellentétes, a központi terület bír gátló hatással, a perifériális részek pedig gerjesztik a ganglionok tüzelését.
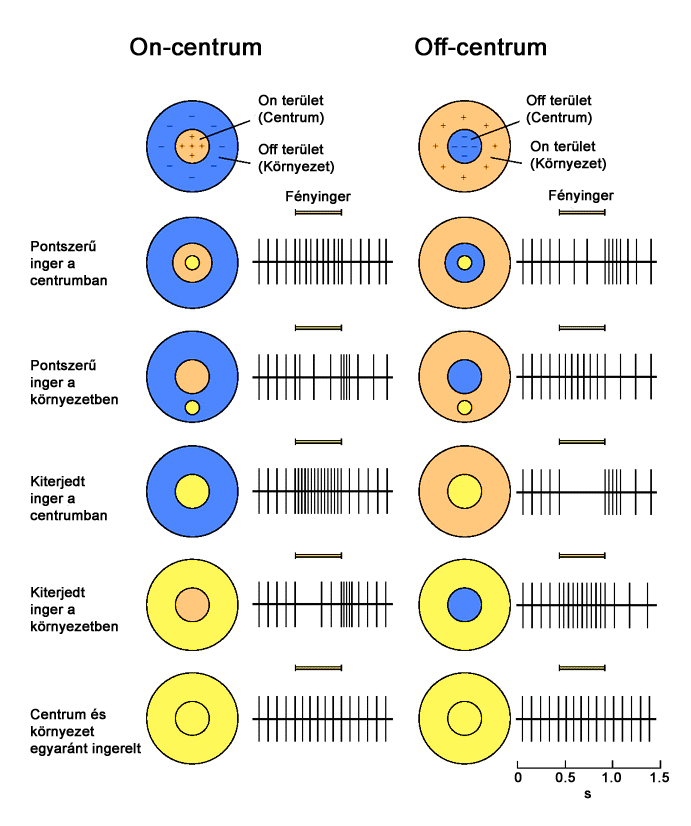
A foveolában található csapok kapcsolódásai jellemzően a direkt utat követik. A receptor mezők mérete itt a legkisebb, akár egyes csapok is rendelkezhetnek külön kapcsolódási útvonallal, amíg a retina perifériális részei felé haladva a receptorok egyre nagyobb méretű receptív mezőket alkotnak. Ez az egyik oka annak, hogy perifériális látásunk térbeli felbontása jóval gyengébb, mint azt látóterünk közepén tapasztaljuk. Ahogy a pálcikák alkotta mezők esetében, úgy a nagyobb méretű csap receptor mezők kialakításában az egyes receptorokból érkező jelek összefűzésével, esetenként a jelfolyam módosításával az amakrin és horizontális sejtek is szerepet játszanak.
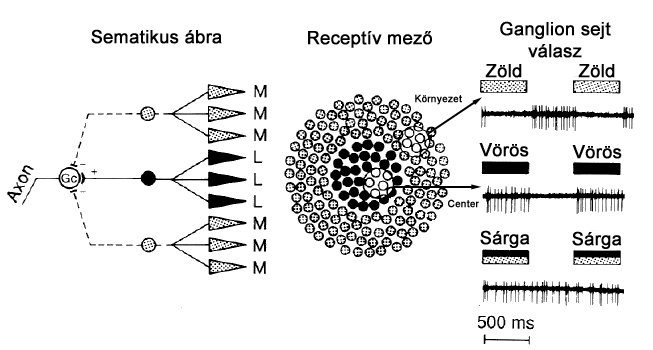
A ganglion sejtek alkotják a látókéreg előtti utolsó állomást, így ezen sejtek akciós potenciáljai képezik a retina kimeneti jelét. Ganglion sejtekből négyféle típust különböztetünk meg: PC (vagy P –parvocelluláris), MC (vagy M – magnocelluláris), KC (vagy K – koniocelluláris) és ipRGC (Photosensitive, azaz fényérzékeny) típust. Az PC és MC ganglion sejtek centrális és perifériális részből álló receptormezővel rendelkeznek, előbbiekhez kisebb méretű, utóbbiakhoz nagyobb kiterjedésű receptor mezők tartoznak, és amíg a PC ganglion sejtek színlátásunk alapjait képezik, és csak nagyon csekély mértékben érzékenyek a kontrasztváltozásokra, az MC típusúak nem játszanak fontos szerepet a színérzékelésben. A KC típusú sejtek a többi ganglion típushoz képest kisméretűek, receptív mezejük csak centrális részt tartalmaz, amely kék csapokhoz kapcsolódva mindig on-, vörös vagy zöld csaphoz csatlakozva pedig mindig off-típusú. Szerepük teljes mértékben még nem tisztázott, de a kontrasztérzékelésben van funkciójuk. Az ipRGC típusú ganglion sejtek receptor mezeje sokkal heterogénebb, csapokat és pálcikákat egyaránt tartalmaz, és az általuk továbbított ingerületek nem a vizuális ingerek kialakításában játszanak szerepet, hanem a fény nonvizuális hatásainak formálásáért felelnek.
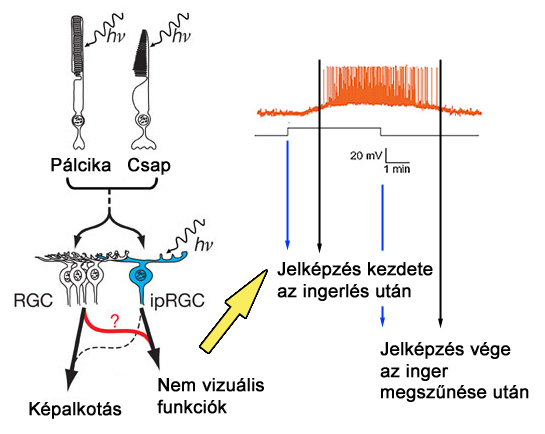
A legújabb kutatások szerint az ipRGC ganglionok önmagukban is fényérzékenyek, tartalmaznak ugyanis opszin molekulát, az úgynevezett melanopszint. Ezen vegyület lebomlásával önmagában is eredményezhet jelképzést a sejt kimenetén, azonban a teljes hatásmechanizmus működésében a ganglion sejt receptor mezejében elhelyezkedő csapoknak és pálcikáknak is szerepük van. Az ipRGC ganglionok működésére jellemző, hogy lassan reagálnak a beérkező ingerekre, valamint az ingerek megszűnésére is (2.11. ábra). Az ipRGC ganglionok száma elenyésző a többi ganglion típushoz mérten, eloszlásuk a retinán nagyjából egyenletes.
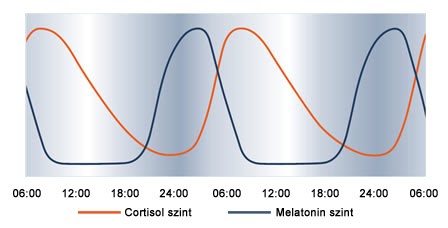
Működésük elsősorban a cirkádián ritmusra (2.12. ábra), vagyis a napszakokkal változó életritmusunkra van hatással, leginkább a melatonin hormon vérbe való kiömlésének elősegítésével és gátlásával. A melatonin mennyisége határozza meg éberségi szintünket - ha ezen hormon szintje magas a véráramban, szervezetünk pihenő üzemmódba kapcsol, elálmosodunk és végül elalszunk. A cirkádián ritmus számos életfunkció váltakozását foglalja magában. Tartalmazza a pulzusszám, vérnyomás és testhőmérsékletet változását, valamint a melatoninon kívül egyéb hormonok, például a cortisol szintjét is. A melanopszin molekula színképi érzékenységének maximuma a látható tartomány kék és ibolya szegmensébe tehető (2.13. ábra). Amikor az ipRGC ganglion sejtet és receptor mezejét olyan spektrális teljesítmény eloszlású fény ingerli, amely nagy mennyiségben tartalmaz kék komponenst, a melatonin hormon termelődése és kiömlése gátolt. Ha az ipRGC ganglionok ingerlése megszűnik, a vér melatonin szintje megemelkedik.
A vizuális érszékelésben résztvevő pálcikák legtöbbje direkt kapcsolattal rendelkező, nagy kiterjedésű receptormezőt alkot. A pálcikák perifériális elhelyezkedésén túl ez okozza éjjeli látásunk rosszabb felbontóképességét. Cserébe a csapokhoz képest jóval érzékenyebb receptorok jeltovábbítása is gyorsabb a nappali látás által igénybevett csatornák jelterjedési sebességéhez képest. Szürkületi látáskor a pálcikák jelei réskapcsolatokon keresztül a csapoknak adódnak át, lehetővé téve ezzel a kétféle receptor együttes működését olyan megvilágítási körülmények között, amely ezt indokolttá teszi – a csapoknak már túl kicsi, a pálcikáknak még túl nagy megvilágítási szint. Sötétben a réskapcsolatok záródnak, a pálcikák jeltovábbítása pedig a bipoláris sejteken keresztül folyik tovább. A szürkületi, vagy más néven mezopos látás különös fontossággal bír járműoptikai alkalmazások esetén, ezért annak sajátosságaival a későbbiekben külön alfejezetben foglalkozunk.
2.2. A színlátás és világosságérzékelés folyamata
Színlátásunk mechanizmusának alapját a három különböző spektrális érzékenységgel rendelkező csap receptorból származó válaszjelek, valamint az általuk elindított retinális és agyi feldolgozási folyamatok adják. Az előbbiekben ismertettük a fotoreceptorok és a hozzájuk kapcsolódó további retinális neuronok működését, ezen fejezetben pedig a színérzékelés rendszerszintű összefüggéseivel foglalkozunk.
A trikromázia, vagyis a három eltérő érzékenységű fotoreceptor együttes működésének elmélete már jóval azelőtt alakot öltött, mintsem a csap receptorok három típusának fiziológiai igazolása megtörtént volna. Ennek alapját az a megfigyelés képezte, hogy három különböző, egymástól független alapszín additív keverékéből bármelyikszíninger létrehozható - az alapszínek függetlenségének kritériuma azt jelenti, hogy egyik alapszín sem lehet előállítható a másik kettő keverékeként. A színlátás trikromatikus szemléletű kutatásának úttörői, Young és Helmholz tehát pusztán elméleti alapon jutottak később helyesnek bizonyuló fiziológiai következtetésekre.
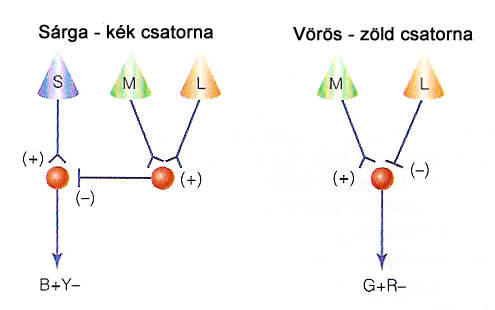
Hasonlóan elvi gyökerekkel rendelkezik a háromszín teóriát kiegészítő opponencia elmélet, vagy antagonisztikus szemléletmód, amely Hering nevéhez köthető. Az opponencia elmélet kiindulási alapja az a felismerés volt, hogy az alapszíneknek tekintett színingereknek vannak olyan kombinációi, amelyek logikailag elképzelhetőek, mégsem társul hozzájuk önálló színfogalom. Ennek megfelelően nem érzékelünk és nevezünk meg vöröses-zöld, vagy kékes-sárga színingereket, ellentétben a sárgás-zöld és kékes-zöld (vagy türkiz) illetve a sárgás-vörös (narancssárga) vagy kékes-vörös (bíbor) ingerekkel, amelyek minden épszínlátó számára ismeretesek. Ebből az a következtetés vonható le, hogy a „köztes színingerekkel” nem rendelkező színpárok úgynevezett alap színpárok, a színingerekre vonatkozó információ pedig ezeknek megfelelő két kromatikus csatornán az alapszínpárok különbségértékeiként kódolódnak. Harmadik opponens csatornaként számon tartunk egy akromatikus, azaz színingert nem, csak intenzitás értékeket kódoló csatornajelet is.
Elsőre úgy tűnhet, hogy a trikromácia és az opponencia elmélete nehezen egyeztethető össze, ennek okán a pontos fiziológiai háttér megismeréséig a két elméletet egymással szembenállónak tartották. A 20. század második felében elvégzett pszichofizikai kísérletek azonban igazolták, hogy a két elmélet a színlátás két különböző szintjének működését vázolja. A primer szint a három eltérő színképi érzékenységgel rendelkező csap receptor válaszjele, amelyek további retinális feldolgozási mechanizmusokon keresztül alakulnak az opponencia elmélet által leírt csatornajelekké (2.14. ábra).
Az akromatikus csatornajel, a környezet objektumainak világosság jellemzőivel kapcsolatban kódol információt. Az emberi szem világosságérzékelésének spektrális vizsgálatai során kimutatták, hogy a nappali látásérzékelésünk hullámhosszfüggő hatékonyságát leíró függvény (a V(λ) függvény, lásd később) jól közelíthető a vörös-érzékeny (L) és zöld-érzékeny (M) csapok érzékenységi karakterisztikáinak súlyozott összegével. Ez az összegzés az erre specializálódott ganglion sejteken keresztül történik. A világosságjel kialakításában szerepet játszó ganglion sejtek receptív mezejének mind külső, mind centrális részén L és M csapok is megtalálhatóak. Ezek jellemzően nagy kiterjedésű, széles laterális kapcsolatrendszerrel bíró mezők, melyek közt mind On- mind Off- centrum típusúak is megtalálhatóak. On-centrum esetében a receptív mező középpontjának ingerlésére nő meg a ganglion sejtek tüzelési frekvenciája, így ez a mechanizmus a sötét háttér előtt megjelenő világos objektumok érzékelését végzi. Off-centrum esetben a hatás pont az előző fordítottja, a környezetet alkotó csapok ingerlése gerjesztő, míg a centrumra eső fény gátló hatású a ganglion sejt kimenetére nézve, így a világos háttér előtt megjelenő sötét objektumok idéznek elő magasabb tüzelési frekvenciát. Különbség még az On-centrum és Off-centrum mezők között, hogy működésük jellegéből adódóan utóbbiak kontrasztérzékenysége nagyobb. Egyenletesen világos környezet esetében mindkét mezőkialakítás kimenete átlagos frekvenciával tüzel, hiszen a gerjesztő és gátló mezők egyszerre ingereltek, egyenletesen sötét háttér esetén pedig egyik esetben sincs jelképzés. Az akromatikus csatornajelet kialakító mechanizmus hatására jön létre a laterális gátlásnak nevezett folyamat, amelynek segítségével a receptor mezők szerkezetéből adódó következmények jól szemléltethetőek. Az ilyen és ehhez hasonló retinális feldolgozási folyamatok esetenként optikai csalódásokon keresztül érhetőek tetten. A laterális gátlás működésének szemléltetésére az úgynevezett Hermann rács alkalmas, ahol nagyobb sötét tartományok között keskeny világos sávok futnak (2.15. ábra). Az ábrát vizsgálva feltűnik, hogy a látómezőnk perifériális részén a csomópontokban sötét foltokat érzékelünk, holott tudjuk, hogy a világos sávok kialakítása homogén. Az is feltűnik, hogy látómezőnk centrális részén a jelenség nem megfigyelhető. A fekete foltok kialakulásának magyarázata az, hogy amikor a szomszédos sötét mezők oldalai közti világos sávok képe On-centrum típusú receptív mezőre esik a retinán, a mezőhöz kapcsolódó ganglion sejt erős gerjesztést kap, hiszen a gátló területekre csak vertikális vagy horizontális irányban esik fény, így a gátló mező területének nagy része nem kap gerjesztést. A csomópontokban, azaz a sötét mezők csúcsainál viszont a perifériális gátló mezőterület ingerlése az itt horizontális és vertikális irányban is jelenlévő csíkozatmiatt kétszeres az élek mentén kialakuló körülményekhez képest, így az agy azt az információt kapja, hogy a csúcsok közti terület sötétebb. Ez egyfajta kontrasztkiemelő hatást eredményez, melynek következtében a kiterjedtebb sötét határral rendelkező világos képrészek intenzívebbnek tűnnek.

Az akromatikus csatorna jeltovábbítása vastagabb, magnocelluláris idegpályákon, MC típusú ganglion sejtek közreműködésével történik, így a jelfolyam gyorsabb, de jóval kisebb térbeli felbontású, azaz nem képes a vizuális ingerek finom részleteinek érzékelésére. Elmondható továbbá, hogy a retina centrális részére nem jellemző ez a fajta mező-szerkezet - ez a (2.15. ábra) ábrán is tetten érhető, hiszen a látómezőnk közepére pozícionált csomópont esetében nem figyelhető meg a periférián jól érzékelhető gátló hatás. Ennek oka, hogy a retina centrális részén a receptív mezők finomabb szerkezetűek. Megemlítendő még, hogy egyes források szerint az S csap is részt vesz az akromatikus csatornajel képzésében, de hatása a végső jelalakra igen csekély, gyakorlati szempontból elhanyagolható.
A kromatikus, vagyis színi információkat is kódoló csatornák közül a vörös-zöld opponenciát alkotó mechanizmus működése nagyon hasonló az akromatikus csatorna képzéséhez, és a foveális területen a színérzékelés mellett a nagyfelbontású kontrasztérzékelést is az elsősorban a vörös-zöld kromatikus csatorna formálását végző receptorok szolgálják ki. Eltérés a receptív mezők szerkezetében, valamint a jeltovábbítást végző idegpályákban és a hozzájuk kapcsolódó ganglion sejtek típusában van. A vörös-zöld opponens jel képzésében részt vevő receptor mezőket is L és M csapok alkotják, a mezők szerkezete azonban jóval rendezettebb, mint az akromatikus jelcsatorna esetében. Itt ugyanis a mezők centrális és perifériális része kizárólag egy-egy típust tartalmaz. Ha a centrumban L csapok helyezkednek el, a környezetben M típusúak, és fordítva. L típusú csapok alkotta centrum esetén, ha a beérkező fény vörös, a ganglion sejt tüzelési frekvenciája nagyobb lesz. Zöld fény beesése esetén nincs kimenő jel, mert a periférián elhelyezkedő M csapok ebben az esetben gátló hatásúak. Sárga fény esetén a gerjesztés és gátlás azonos mértékű, így a tüzelési frekvencia átlagos lesz (2.16. ábra). Ennek megfelelően elmondható, hogy a ganglion sejt kimenetén az akciós potenciálok tüzelési frekvenciája a beeső fény vörös-zöld arányával megegyező nagyságú lesz. M típusú centrummal rendelkező receptív mezők esetében a mechanizmus működése ugyanilyen, csak fordított előjelű. A kromatikus csatornajeleket képző ganglion sejtek PC típusúak, elhelyezkedésük túlnyomó többségben a foveolára koncentrálódik. A receptív mezők szerkezete jóval finomabb, mint az akromatikus jeleket képző mezőké, akár egyetlen direkt kapcsolattal rendelkező csap is alkothatja a mező centrális részét. Ennek a nagyon finom sejtmintázatnak a kialakításához a vaskos idegpályák nem megfelelőek, így a jeltovábbítás nem a már ismert magnocelluláris pályákon, hanem a sokkal vékonyabb axonok (idegsejt nyúlványok) alkotta parvocelluláris idegpályákon történik. Ez lehetővé teszi a vizuális ingerek finomabb részleteinek feldolgozását is.
A sárga-kék kromatikus csatornajel képzése során a mechanizmus működése a fent vázoltakhoz hasonlóan történik, azzal a különbséggel, hogy itt a retinán jóval ritkábban fellelhető kék érzékeny S csapok is szerephez jutnak. A kapcsolódó ganglion sejtek számára ezen receptorok szolgáltatják a kék fény beesése esetén szükséges gerjesztést, a gátló hatást pedig a receptív mező másik részén található M és L csapok alkotta receptor köteg váltja ki. A sárga ingert nem külön csaptípus, hanem az M és L csapok együttes jelenléte biztosítja a receptor mező gátló területein, amelynek hatására a gátlás mértéke ezen csapok együttes ingerlésének mértékével, azaz az L és M csapok jelének összegeként leírható sárga színinger mértékével lesz arányos. A sárga-kék opponens párt képző receptor mezők felépítése a másik kromatikus csatornával megegyező módon finomszerkezetű, a jeltovábbítás itt is a vékonyabb parvocelluláris idegpályákon keresztül történik.
Az így kialakult kromatikus és akromatikus csatornajelek ezután a látóidegen keresztül az agyba továbbítódnak. A rendezett kötegben futó látóidegpályák mentén még az agy releváns részének elérése előtt megkezdődik a jelek feldolgozása, a kép már itt élekre, formákra, tónusokra bomlik, majd a megfelelő axonok az agy tarkó felöli területén megtalálható látókéregbe, más néven cortexbe továbbítják a jeleket. A cortex egyes részeiben történik a vizuális ingerek végső értelmezése, itt alakulnak ki a tudatunkban, emlékeinkben megjelenő képek.
2.3. Látórendszerünk adaptációs mechanizmusai
Látórendszerünk alapvetően kétféle adaptációs mechanizmussal rendelkezik, melyek kialakításában több eltérő funkcionalitással rendelkező folyamat is szerepet játszik. Az összetettebb mechanizmus mindenképpen a világos-sötét illetve a sötét-világos adaptáció többlépcsős folyamata, de a színi adaptáció relatíve egyszerűbb mechanizmusa is komoly hatást fejt ki látásérzékelésünk egészére.
2.3.1. Sötét és világos adaptáció
Ha a sötét-világos arányt, azaz a környezet fénysűrűség értékét, mint környezeti változót vizsgáljuk, elmondható, hogy az emberi látás rendkívül nagy intenzitástartomány átfogására képes. Pusztán néhány foton elegendő ahhoz, hogy jelfolyamot indítson meg a retina különböző rétegei között, és a meginduló inger vizuális észleletté alakuljon. Ugyanakkor nyáron, a déli napfényben retinánkra záporozó milliárdnyi foton által keltett jelek feldolgozása sem jelent akadályt látórendszerünknek, mi több, ilyen körülmények között is csaknem olyan komfortosan érezzük magunkat, mint alacsony vagy általános fénysűrűségű környezetben. Az említett végletek között az észlelt fénysűrűségben és a környezetben található felületek megvilágítottságát tekintve - figyelemre méltó - 6-8 nagyságrend eltérés van. Jelenleg a műszaki területeken alkalmazott szenzorok meg sem közelítik ezt a dinamika-tartományt.
Látórendszerünk erre a figyelemreméltó teljesítményre több különböző mechanizmus együttes működésével, illetve eltérő körülmények között más-más mechanizmusokat működtetve képes. A fényviszonyok megváltozására adott első reakció a pupillareflex. A retinára beérkező fény intenzitásának növekedésével a szivárványhártya közepén található pupillanyílás mind jobban összeszűkül, csökkentve ezzel a szem belső részébe jutó fény mennyiségét. A pupillareflex vezérléséért a korábban leírtak szerint az ipRGC típusú, azaz fényérzékeny ganglion sejtek és a hozzájuk kapcsolódó fotoreceptorok, valamint a jeleiket feldolgozó neurális folyamatok felelősek. Érdemes megjegyezni, hogy a fényérzékeny ganglion sejtek önmagukban, a hozzájuk kapcsolódó receptorok jelei nélkül is képesek a pupillareflex bizonyos mértékű kivezérlésére. Ugyanez fordítva is elmondható, a melanopszin molekulák eltávolításával, ezzel a W ganglion sejt fényérzékenységének megszüntetésével, pusztán a hozzá kapcsolódó receptorok ingereivel is csökevényes pupillareflex figyelhető meg. A teljes funkcionalitását azonban csak a két receptortípus együttes működése biztosítja. A pupilla összehúzódásának sebessége eltérő hullámhosszú ingerlésre nem állandó. A rövidebb hullámhosszú sugárzás jóval gyorsabb összehúzódásra készteti a szivárványhártyát, mint a hosszabb hullámhosszú. Ennek magyarázata a melanopszin molekulák színképi érzékenységében keresendő, amelynek maximuma a rövidebb hullámhosszú, kék tartományra tehető. Fiziológiai szempontból azért indokolt a rövidebb hullámhosszakra való gyorsabb reakció, mert ezen a spektrumtartományon nagyobb energiájú a sugárzás, vagyis az rövidebb idő alatt fejthet ki káros hatást.
A pupillareflexen túl látórendszerünk egyéb, jóval összetettebb mechanizmusokkal is válaszol a környezeti fénymennyiség megváltozására. A nappali látás során működő csapok a fénymennyiség csökkenésével inaktív állapotba kerülnek, és a jóval érzékenyebb pálcikák aktivizálódnak, ezzel látórendszerünk éjjeli üzemmódba vált. A két mechanizmus közötti átváltás nem diszkrét átmenettel, hanem egy bonyolult működésű köztes folyamaton, a mezopos látásmechanizmuson keresztül történik, amikor mindkét receptortípus működésben van. Ezen mechanizmus járműoptikai szempontból vett fontossága miatt külön alfejezetben részletesebben foglalkozunk. A sötét-világos és világos-sötét adaptáció (a továbbiakban világos(ra), illetve sötét(re) adaptáció) dinamikája között akadnak eltérések, ezért a két folyamat különböző módszerekkel vizsgálható.
A sötét adaptáció vizsgálatához a tesztalanyok egy sötétszobában foglalnak helyet, és megadott időközönként be kell állítaniuk egy tesztfelület fénysűrűségét nulláról, a már éppen észlelhető szintre. A vázolt mérés eredményeként kapott görbéket mutatja a 2.17. ábra.
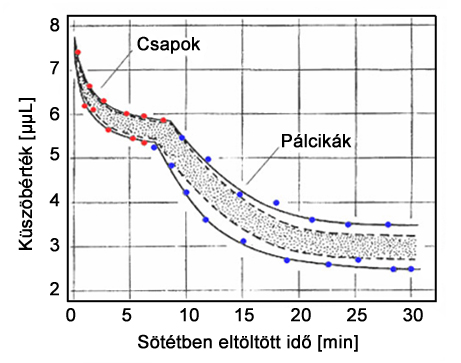
A sötét adaptációs görbéket vizsgálva jól elkülöníthető két hasonló karakterisztikájú, de különálló görbeszakasz. Az adaptációs idő elején magasabb fénysűrűség érték beállítása szükséges a tesztfelület észleléséhez. A fénysűrűség küszöbértéke egy darabig rohamos ütemben csökken, majd a görbe fokozatosan ellaposodik. Ez utóbbi/előbbi görbeszakasz a csapok adaptációs mechanizmusát jeleníti meg, így az ellaposodó görbeszakasz a csapok által érzékelhető legalacsonyabb küszöbértéket mutatja. Ha csak csap típusú receptor sejtekkel rendelkeznénk, ennél alacsonyabb fénysűrűség értékeket nem lennénk képesek érzékelni. Azonban a csapok érzékenységi küszöbét elérve működésbe lépnek a sokkal érzékenyebb pálcikák. A pálcikák sötét adaptációs görbéi alkotják a 2.17. ábra által bemutatott adaptációs karakterisztika második, az első szakasztól jól elkülöníthető részét. Ezek a görbék jellegüket tekintve nagyon hasonlóak a csapok esetében tapasztalhatóakhoz, azonban küszöbértéküket jóval alacsonyabb fénysűrűség szinteken érik el. A 2.17. ábragörbéin az is megfigyelhető, hogy a pálcikák érzékenységi küszöbértékének eléréséhez több sötétben töltött idő eltelte szükséges, mint a csapok esetében.
A sötét adaptációs görbék általános megadáskor görbesereget vagy tartományokat, nem pedig individuális görbéket ábrázolunk. Ennek oka, hogy a görbe pontos alakját számos tényező befolyásolja. A vizsgálat előtti elő-adaptációs körülmények, úgymint a pre-adaptációs környezetben eltöltött idő, és a környezet átlagos megvilágítottsága jelentős mértékben megváltoztatják az adaptációs görbék lefutását. Mindezeken túl a görbealakra hatással van még a tesztfelület megvilágítására használt fény spektrális teljesítmény eloszlása, a csapok és pálcikák heterogén retinális eloszlása miatt a teszt során ingerelt retinatartomány pozíciója, valamint hasonló okokból a tesztfelület átmérője is. Az adaptációs mechanizmusokat befolyásolja még a vakítás jelensége is. Vakítás akkor lép fel, ha a retinára eső fény hatására lebomló fotopigmentek száma a receptorokban nagyobb, mint amennyi azonos időegység alatt újratermelődni képes. Amennyiben a pre-adaptációs környezet fénysűrűsége elég nagy, és a vizsgált személyek által itt töltött idő megfelelő hosszúságú ahhoz, hogy a fotopigmentek termelődése és lebomlása közötti egyensúly megbomoljon vagy ne legyen képes beállni a receptorokban, a hatás befolyással lesz a sötét adaptációs görbék alakjára.
Fontos megjegyezni, hogy a vakítás hatása nem lineáris, hanem logaritmikus, valamint a csapok sötétadaptációjára kevésbé van hatással, mint a pálcikákéra. Az receptorokban megtalálható opszin molekulák mennyisége retinális denzitometriával mérhető, ahol a retináról visszaverődő fény spektrális teljesítmény eloszlásának és abszolút intenzitásának elemzésével állapítható meg a lebomlott fotopigment mennyiség. A módszer alapját az az egyszerű jelenség adja, hogy a pigmentek bomlásukhoz fotonokat abszorbeálnak, így minél magasabb a fotoreceptorokban az opszinok mennyisége, annál kevesebb visszavert fény mérhető egyes hullámhossz tartományokon. A pálcikákban található rodopszin 1%-ának lebomlása hozzávetőlegesen 10 egységgel csökkenti az adaptáció elején tapasztalt kezdeti érzékenységet. Ennek megfelelően a rodopszin molekulák 50%-ának lebomlása 10 logaritmikus egységgel csökkenti a kezdeti érzékenységet, ám a csapok esetében ugyanennyi pigment lebomlása csak másfél logaritmikus egységgel csökkenő érzékenységet eredményez. Sötétadaptáció alatt tehát a csapokban és a pálcikákban található fényérzékeny pigmentmolekulák lebomlásának és termelődésének egyensúlyi állapotba kerülését értjük, vagyis a sötét adaptációs görbék a látásérzékeléshez szükséges idegi jeleket lebomlásukkal elindító pigment molekulák sokaságának felépülési dinamikáját mutatják.
Világos adaptáció esetén látórendszerünknek a pre-adaptációs környezethez képest magasabb fénysűrűségű háttérhez kell alkalmazkodni, hogy a háttér előtt megjelenő eltérő fénysűrűségű céltárgy detektálható legyen. A világos adaptáció lefolyása az úgynevezett küszöbérték növekményméréssel vizsgálható (2.18. ábra). Ennél a vizsgálati módszernél a vizsgálati személy egy adott fénysűrűségű háttér közepén található célterület fénysűrűségét változtathatja egészen addig, amíg a céltárgy vizuálisan elkülöníthetővé válik a háttértől. Ennek megfelelően a mérés során a háttér fénysűrűsége a független, a céltárgy fénysűrűsége pedig a függő változó.
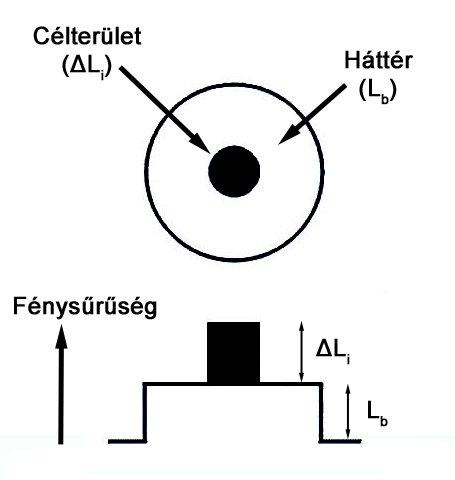
A 2.18. ábra által mutatott mérési elrendezés lényege, hogy a vizsgálat a háttér különböző fénysűrűség értékeinek beállításával is megismételhető, így a háttér fénysűrűsége és az észlelési küszöbérték közötti összefüggés grafikon formájában felrajzolható - ez az úgynevezett TVI („threshold-versus-intensity”) görbe. Akárcsak sötét adaptáció esetén, a világos adaptáció működését szemléltető görbe is két részre bontható (2.19. ábra). A háttér alacsonyabb fénysűrűség értékeihez tartozó görbeszakasz a pálcikákhoz tartozó küszöbérték növekményt ábrázolja, a magasabb háttérfénysűrűségek esetén pedig az első görbeszakasztól jól elkülöníthetően látszik a csapok világos adaptációs mechanizmusának lefutása. A görbék ábrázolása a világos adaptáció jellemzésekor logaritmikus skálán történik, lefutásukat pedig a sötét adaptációs görbék esetén fentebb említett külső tényezők befolyásolják.
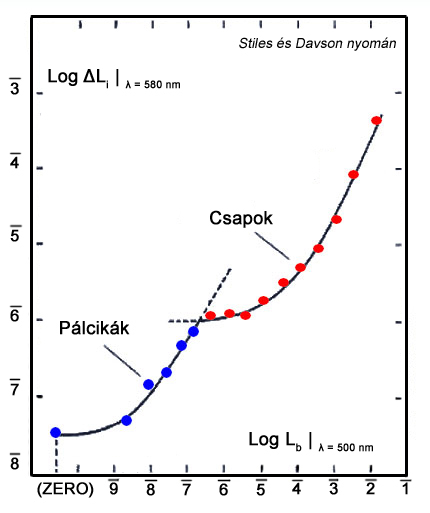
Mivel a világos adaptációs görbék mérésekor alkalmazott küszöbérték növekmény függő és független változója is könnyen kontrolálható, ez a módszer a sötét adaptációs méréshez képest jobb lehetőséget ad az adaptációs mechanizmus összetettebb vizsgálatára, hiszen ott a független változó – a sötétben eltöltött idő – természeténél fogva nehézkessé teszi a tesztek precízen kontrolált kivitelezését. A küszöbérték növekmény vizsgálat során a háttér és a célterület spektrális teljesítmény eloszlásának megfelelő megválasztásával a pálcikák alkotta rendszer izoláltan gerjeszthető, így a pálcikák működéséhez tartozó görbék pontos karakterisztikája, finomszerkezete is megfigyelhető (Aguilar és Stiles, 1954). Ehhez a hátteret vörös színűre kell választani, így a célterület környezetének jellemzően hosszú hullámhosszú sugárzása a vakítás mechanizmusa miatt - vagyis a receptorokban található fotopigmentek nagy részének lebontásával – telítésbe viszi a csapok alkotta vizuális rendszert, amelynek következtében megfelelően megválasztott zöld színű célterülettel csak a pálcikák alkotta válaszjelek lesznek befolyással a tesztalanyban keltett végső észleletre, így a mérés eredményére. Az ezzel a módszerrel mért, pálcikákhoz tartozó küszöbérték növekmény görbék jól láthatóan négy részre bonthatóak (2.20. ábra).
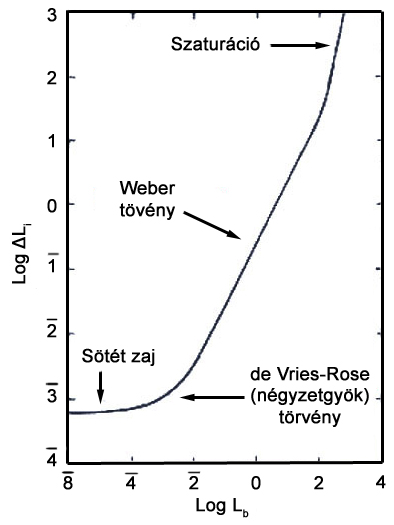
Az első szakasz az úgynevezett „sötét zaj”, amely a viszonylagos instabilitásuk miatt gerjesztés nélkül, vagy termikus hatások eredményeként lebomló opszinmulekulák által indukált jel, valamint a jelképzés neurális rendszerének más elemeiből származó zajok szuperpozíciójaként adódik.
Ezt követi a négyzetgyök törvény, vagy más néven de Vries-Rose törvény által jellemzett szakasz. Ez a szakasz képez átmenetet a sötét zaj tartománya, és a Weber törvény által jellemzett tartomány között. A látórendszer működésének vizsgálatához annak viselkedését legtöbbször egy teoretikus rendszerhez, az ideális fénydetektor működéséhez hasonlítjuk. Az ideális detektort definíciója szerint minden ráeső fotont elnyel, így érzékelési képességeit kizárólag a fényforrás kvantumfluktuációja, vagyis a fotonok keletkezésének véletlenszerűsége határolja be. Rose (1948) szerint ennek megfelelően a látórendszer érzékenységének is egy adott tartományon erős összefüggést kell mutatnia a háttér kvantumfluktuációjával, így arányosnak kell lennie a háttér fénysűrűségének négyzetgyökével. A világos adaptációs görbe második szakasza nem felel meg tökéletesen ennek a szabályszerűségnek, de jellegét tekintve egyezést mutat, a csekély eltérést pedig a következő görbeszakaszba történő tranziens átfutás okozza.
A harmadik, egyben talán a legfontosabb szakasz a korábban már említetteknek megfelelően a Weber törvény által jellemezett tartomány, illetve az a szakasz, amelyre a törvény igaznak mondható. A Weber törvény szerint a háttér fénysűrűségének és a célterület küszöbérték növekményének aránya állandó. Ez a törvényszerűség látórendszerünk egyik alapvető jellemzőjére világít rá. A mindennapi életben leggyakoribb vizuális feladat adott tárgyak elkülönítése különböző hátterektől. A megkülönböztetéshez a vizsgált tárgyaknak minden esetben rendelkezniük kell valamekkora kontraszttal a háttérhez képest, ezen a kontrasztnak pedig függetlennek kell lennie a megvilágítás intenzitásától. A harmadik görbetartomány az, ahol ez a függetlenség fennáll, így elmondható, hogy látórendszerünk ezen a tartományon működik rendeltetésszerűen. Ezen a szakaszon definiálható az úgynevezett Weber konstans, amely a háttér és a tőle már éppen megkülönböztethető céltárgy közötti kontrasztarány. Ez az arány a pálcikákra 0.14 (Cornsweet, 1970), a vörös és zöld érzékeny csapokra rendre 0.02 és 0.03 (Davson, 1990), a kék érzékeny csapokra pedig 0.09 (Stiles, 1959) értékekre adódik.
A negyedik görbetartomány a szaturációs szakasz. Itt a háttér nagy fénysűrűség értékei telítésbe viszik a gerjesztett pálcikákat, azaz a bennük található fényérzékeny rodpopszin gyorsabban bomlik le, mint ahogy újratermelődni képes. Ezen körülmények között a világos adaptációs görbe pálcikák jellemezte szakaszának meredeksége emelkedni kezd, majd működésbe lépnek a csapok, ezzel pedig a világos adaptáció következő szakasza. A csapok világos adaptációjának működése a pálcikákkal megegyező módon zajlik. Különbség mindössze az intenzitási szintekben, illetve a Weber törvény által jellemzett lineáris szakasz meredekségében tapasztalható.
2.3.2. Színi adaptáció és más színlátást befolyásoló mechanizmusok
A sötét és világos adaptációval látórendszerünk a környezet fénysűrűségének megváltozását követi. Van azonban egy másik, ettől eltérő adaptációs mechanizmus is, amely a környezetet megvilágító sugárzás spektrális teljesítmény eloszlásának globális megváltozására, valamint a vizsgált tárgy környezeti viszonyaira reagál - ez az úgynevezett színi adaptáció, színkonstancia vagy von Kries-féle adaptáció.
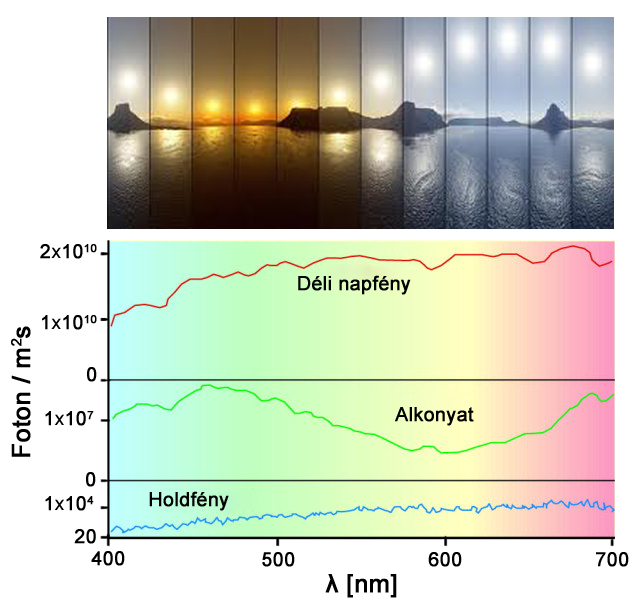
Ahhoz, hogy a különböző hátterek előtt ne csak megkülönböztetni, hanem felismerni is képesek legyünk a tárgyakat, a kontrasztérzékelés intenzitásfüggetlenségéhez hasonlóan különböző környezeti körülmények esetén szükség van a színérzékelés bizonyos mértékű függetlenségére is. Egy papírlapot a déli napsugárzás sokkal magasabb színhőmérsékletű, azaz kékesebb fényénél is ugyanúgy fehérnek érzékelünk, mint a felkelő vagy lemenő Nap sugarai által megvilágítva, amikor is a légkör abszorpciós hatásának növekedésével – nagyobb légtömegen halad keresztül a sugárzás – a környezet vörösesebbé válik (2.21. ábra).
Látórendszerünk ezen alkalmazkodási képességét nevezzük színi adaptációnak. A mechanizmus alapját a vörös-, zöld- és kék-érzékeny fotopigmentek lebomlása és termelődése közötti egyensúly beállása jelenti. A korábban felhozott fehér papírlapos példán keresztül a jelenség könnyen megérthető. Egy céltárgy színét akkor érzékeljük neutrálisnak, azaz fehérnek, ha az arról reflektált fény mind a három különböző típusú csap receptort hozzávetőlegesen azonos mértékben ingerli. Ha alapesetben feltételezzük, hogy a céltárgyat megvilágító fény spektrális teljesítmény eloszlása ekvienergikus – vagyis minden hullámhosszon azonos intenzitású –, a csapreceptorok érzékenységének maximum értékei úgy fognak alakulni egymáshoz képest, hogy a görbék alatti terület egységnyi értéket vegyen fel, azaz a három eltérő érzékenységű receptor által adott válaszjel egyenlő legyen (2.22. ábra).
Abban az esetben, ha a céltárgy megvilágításának spektrális teljesítmény eloszlása megváltozik, a receptorok a korábbitól eltérő mértékben ingerlődnek – ilyen módon pedig a korábbiakban neutrális fehér papírlapot valamilyen színezettel rendelkezőnek érzékeljük. Ilyenkor azonban a receptorokban található pigment molekulák bomlása és termelődése közötti egyensúly is megbomlik. Ez az egyensúly azonban idővel – néhány perc elteltével- helyreáll, és így a fehér papír újra színezet nélküli, neutrális észleletet kelt. A teljes mechanizmus működése ennél jóval összetettebb, a retinális ingerfeldolgozásnál sokkal mélyebb kognitív területeket is érint, azonban elmondható, hogy a színi adaptációs képesség fundamentumait a fényérzékeny pigmentek bomlásának és termelődésének egyensúlya teremti meg. A színi áthangolódás azonban csak bizonyos határok között működik, vagyis neutrális észlelet az észlelő színterében az ekvienergikus környezeti sugárzás hatására beálló neutrális pont meghatározott környezetébe eső színpontokra jöhet létre.
Ha nem neutrális ingerből indulunk ki, hanem egy monokromatikus fényhez keverünk egyre több neutrális hátteret, vagy növeljük a monokromatikus fény spektrális eloszlásának félérték szélességét - azaz a kiinduló színünket tesszük egyre telítetlenebbé -, azt figyelhetjük meg, hogy az észlelt színnek nemcsak a telítettsége, de kis mértékben a színezete is változik (2.23. ábra). Ez az úgynevezett Abney effektus, amely nevét első megfigyelőjéről, Sir William Abneyról kapta. A jelenség érdekessége, hogy eltérő hullámhossz tartományokon más-más irányú eltolódást eredményez. 500 nm alatt a deszaturált inger által keltett észlelet kékesebbnek, 500 nm felett sárgásabbnak tűnik. A telítettség csökkenésével az adott színinger által kiváltott világosság észlelet sem az elvárt módon változik. Azonos fénysűrűség mellett az erősen telített színek világosabb érzetet váltanak ki, mint azok deszaturált árnyalatai. Ez a jelenség az úgynevezett Helmholtz-Kohlrausch effektus, amely alapján elmondható, hogy a kromatikus világosságérzékelés nem követi Weber linearitási törvényét.

A környezet spektrális teljesítmény eloszlásán kívül a környezeti fény intenzitásváltozása is hatással van színlátásunkra, ez az úgynevezett Bezold-Brücke eltolódás. Ha növeljük a céltárgyról visszavert fény intenzitását, az Abney effektussal megegyező hatást tapasztalhatunk, azaz 500 nm alatt kékesebbnek, 500 nm felett sárgásabbnak tűnnek az árnyalatok. Ha azonban a céltárgy megvilágítottsága csökken, a hatás leginkább a vörös-zöld tartományon jelentkezik. Egy vörös virágot szemlélve zöld levelek előtt, először a szirmok veszítenek nagyobb mértékben telítettségükből, ahogy csökken a megvilágítás intenzitása, egészen addig, amíg bele nem olvadnak a még kissé zöldnek tűnő levelek alkotta háttérbe, majd a megvilágítás további csökkenésével végül mind a szirmok mind a levelek elveszítik színezetüket.

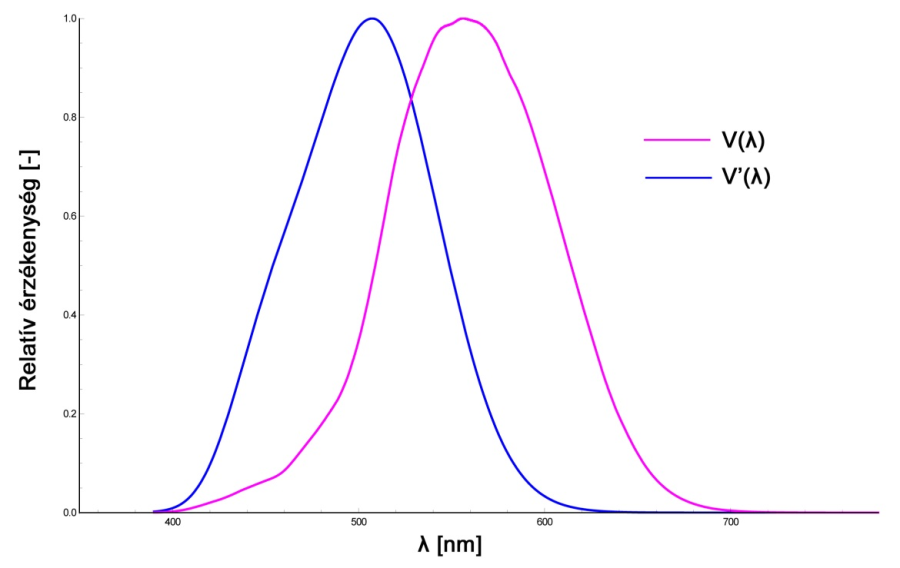
Az úgynevezett Purkinje effektus (2.24. ábra), létrejötte a nappali vagy fotopos fényhatásfok függvény - vagyis a V(λ) görbe - és az éjjeli, más néven szkotopos fényhatásfok függvény – azaz a V’(λ) görbe - közti átmenettel magyarázható. Előbbi esetében a csaplátás a domináns látási mechanizmus, utóbbi esetében pedig a pálcikák dominálta látórendszer működik. A kettő közötti átmenet során látórendszerünk érzékenységi maximuma a rövidebb hullámhosszak felé tolódik - ez az átmeneti tartomány képezi a mezopos látástartományt.
2.3.3. Az emberi látás működése különböző megvilágítási szintek mellett
A korábbiakban már érintőlegesen szó esett a fotopos és szkotopos látásról, valamint a kettő közötti átmenetet képező mezopos látástartományról. Foglaljuk össze röviden, hogy mit is takarnak ezek a fogalmak, és hogyan kapcsolódnak egymáshoz, valamint látórendszerünk működésének más területeihez.
A világosságérzékelés vizsgálatára alkalmazott modern módszer az úgynevezett villogásos fotometria. Az ilyen mérések során látórendszerünk azon sajátossága kerül kihasználásra, hogy a retinális jelképzés során a magasabb neurális szintek felé magnocelluláris idegpályákon közlekedő világosságjel akkor is ad az agy számára értelmezhető jeleket, ha a látómező egyes területeinek gyors változását a parvocelluláris pályákat használó színcsatornák már nem tudják követni.
Ennek kiaknázása méréstechnikai szempontból azért hasznos, mert eltérő színképi összetételű fények világosságának összehasonlítása még rendkívül precíz optikai felépítésű fotométerekkel is nehézkes. Ha azonban a kromatikus és akromatikus jelek terjedési sebessége közötti különbséget kihasználva, adott frekvencián elkezdjük egymással felcserélve „villogtatni” a különböző spektrális teljesítmény eloszlású ingereket, kezdetben azt tapasztaljuk, hogy mind a színezetbeli, mind a világosságbeli pulzálás érzékelhető. A frekvencia növelésével egy adott frekvenciaszintnél - amelynek pontos értéke függ a besugárzás szintjétől, de jellemzően 15-20 Hz környékére tehető – a színezetbeli fluktuáció megszűnik, és csak a világosságszintek közötti vibrálás érzékelhető. A frekvencia további növelésével elérjük az abszolút fúziós frekvenciát, ahol már a világosságértékek közötti különbség sem látható, a két felület megvilágításának átlagértékét érzékeljük.
A köztes szakasz lehetőséget ad arra, hogy a világosságérzékelést közvetlen módon, a színezetbeli eltérések szubjektív kognitív hatásainak kiküszöbölésével vizsgálhassuk. Ha a villogási frekvenciát ilyen értékre választjuk, a két fényforrás által a vizsgálati mező két oldalán keltett megvilágítás relatív értékeinek állításával elérhető a villogás érzetének megszüntetése, amely adott beállítások mellett a mezők világosságának érzékelése közötti egyezést jelenti.
A fenti módszerrel a látómező egyik oldalát heterokromatikus, másik oldalát pedig adott hullámhosszúságú monokromatikus fénnyel megvilágítva, hullámhosszról-hullámhosszra meghatározható az emberi világosságérzékelés színképi karakterisztikája.
A Nemzetközi Világítástechnikai Bizottság (Comission Internationale d‘Éclairage, CIE) 1924-ben rögzítette az átlagos "láthatósági" függvényt, s ezt a visibility szó kezdőbetűje alapján V(λ) függvénynek nevezték. Ahogy arról már korábban is szót ejtettünk, a napjainkban is használt fotometriai rendszer egyik alappillére ez a V(λ)-görbe. A görbe elfogadásakor több eltérő módszerrel és különböző észlelőkkel elvégzett kísérletek eredményeinek összevetésével és közös nevezőre hozásával, valamint a jellemzően 490 és 680 nm közé eső mért tartományokon túl 400 nm-ig és 760 nm-ig történő extrapolálással határozták meg a végső függvényalakot. A görbét az 1924-es ülésen, mint ideiglenes megoldást fogadták el, célul tűzve ki annak későbbi módosítását. A görbék pontosítására csak 1991-ben került sor, módosítások pedig jórészt csak az extrapolált szakaszokon történtek. Az eredeti és a pontosított görbék közötti csekély eltérés jól mutatja az 1924-ben meghatározott függvényalak viszonylagos precizitását.
Az emberi szem érzékenységét így a V(λ) és V’(λ) függvények illusztrálják. A csapok, azaz a nappali látás receptorainak spektrális érzékenységét a V(λ), az éjszakai látás receptorainak (pálcikák) érzékenységét pedig a V’(λ) függvény szemlélteti (2.25. ábra). Ahogyan azt már a retinális jelfeldolgozást taglaló fejezet világosságérzékeléssel kapcsolatos részében említettük, jelenlegi ismereteink szerint a V(λ) görbe képzésében jellemzően vörös és zöld érzékeny csapok alkotta receptormezők vesznek részt. Ennek megfelelően a V(λ) görbe jelenlegi definíciója alapján a Stockman és Sharpe által 2000-ben meghatározott vörös és zöld csapérzékenységi görbék szuperpozíciójaként is megadható.
Három látási mechanizmust különböztetünk meg:
-
Fotopikus vagy fotopos látás:
a V(λ) által illusztrált nappali látás, amely során a szem receptorai közül csak a csapok működnek. Ez körülbelül 3 cd/m2-nél nagyobb fénysűrűség esetén teljesül. Ezt hívjuk fotopos fénysűrűségi tartománynak. Egyezményes felső határa nincs, de 105 cd/m2 fölött káprázási, majd vakítási jelenségek lépnek fel, ilyen körülmények között a fotometria additivitási, proporcionalítási stb. törvényei már nem teljesülnek, így a fotometriai leírás érvényét veszti.
-
Szkotopikus vagy szkotopos látás:
az éjszakai látás, mely során csak a V’(λ) által meghatározott pálcikák működnek, melyek érzékenysége 1 cd/m2 alatt van.
-
Mezopikus vagy mezopos látás:
köztes, úgynevezett szürkületi látás, amikor a csapok és a pálcikák együttesen működnek. A 10-3 cd/m2 és 3 cd/m2 közti fénysűrűség tartományt mezopos tartománynak hívjuk. A fotopos tartománytól kezdve folyamatosan csökken a csapok szerepe, és nő a pálcikák szerepe a látásba. Ennek megfelelően fokozatosan tolódik el a fényérzet színképi érzékenysége a V(λ)-tól a V’(λ) görbe felé. Gyakorlati szerepe miatt (például szürkületi vezetés) megismerésére és vizsgálatára jelenleg is kísérletek folynak, de nemzetközileg még egyik rendszert sem fogadták el (CIE 81-1989). A mezopos fotometria kutatásának főbb állomásaival és jellemzőivel fontossága miatt a következő alfejezetben részletesebben is foglalkozunk.
A fotopostól eltérő szkotopos és mezopos fotometria használata bizonyos területeken és helyzetekben jelentős. Egy nem megfelelően használt rendszer (pl. fotopos a mezopos vagy szkotopos helyett) komoly hibákhoz vezethet. Elég csak a közlekedésre vagy a vezetésre gondolnunk. Az útvilágítás területén az előforduló fénysűrűségi értékek általában a mezopos tartományba esnek.
Ezt egy gyakorlati példa szemlélteti. Ha az útburkolat fénysűrűségét 0,05 cd/m2-nek választjuk meg, úgy a mezopos, vagy szkotopos fotometria rendszerét használva kisnyomású nátrium lámpa (aranysárga fényű) és nagynyomású higanylámpa (kékeszöld fényű) mért fénysűrűségének értéke miként változik meg:
|
Na (cd/m2) |
Hg (cd/m2) |
|
|---|---|---|
|
Fotopos |
0,050 |
0,050 |
|
Mezopos |
0,028 |
0,061 |
|
Szkotopos |
0,010 |
0,070 |
Ezek alapján látható, hogy a nagynyomású Hg lámpa mezopos vagy szkotopos fotometria szerint magasabb fénysűrűségi értékeket mutat, mint a Na lámpa. Ha például arra vagyunk kíváncsiak, hogy egy úton lévő akadályt milyen könnyen és gyorsan veszünk észre, úgy a szem csap-mechanizmusának ingerlését figyelembe kell venni még szkotopos illetve mezopos tartományban is.
2.3.3.1. A mezopos látás sajátosságai
A fenti példa jól mutatja, hogy a járművekben feladatot teljesítő világító berendezések tervezésekor fontos a mezopos látástartomány behatóbb ismerete, valamint a mezopos fotometriára vonatkozó kísérletek és nemzetközi ajánlások áttekintése.
A korábbiakban a világos és a színi adaptációval kapcsolatban is szó esett a pálcikák dominálta, azaz a szkotopos és a csapok dominálta, azaz a fotopos látás közötti átmenetről. A csapok és pálcikák közötti átváltás az úgynevezett mezopos látás tartományán keresztül történik, ahol a normál körülményekhez képest alacsonyabb hatásfokkal, de mindkét receptor típus működik.
A csapok fénysűrűség érzékelésének lineáris szakaszán Abney törvénye érvényesül, amely szerint, ha A és B színingerek egyforma világosságúak, valamint ha C és D színingerek is egyforma világosságúak, akkor A és C, valamint B és D ingerek additív keverékei is egyforma világosságúak. Ez a törvény képezi a nappali fotometria alapját. A világosság adaptációnál leírtak alapján a mezopos fénysűrűség tartományon a pálcikák már szaturációs állapotban vannak, a csapok pedig még nem értik el a Weber törvény által jellemzett lineáris működési tartományt. Ennek következtében az ilyen háttérfénysűrűséggel jellemezhető környezetben látórendszerünk működése nem lineáris, Abney összegezhetőségi törvénye nem teljesül.
Nem szabad azonban megfeledkezni arról sem, hogy az említett összegezhetőségi törvény csak akromatikus fényekre igaz, látórendszerünk pedig nem tisztán a vizsgált tárgy világossága alapján határozza meg annak láthatóságát, hanem a tárgy és környezete közti szín- és világosságkontraszt alapján – a színészlelés pedig, mint arról korábban szó esett, nem független a világosságtól. A mezopos látás tehát igen összetett jelenség, vizsgálata nagyfokú körültekintést igényel.
A mezopos fotometria kutatásának kezdetekor nagyobb hangsúly került a mezopos világosság észlelés vizsgálatára, mert ezen tartományban a pálcikák alkotta receptormezők nagyobb mérete, és a jelentősen jobb felbontású csapok kismértékű ingerlése miatt a vizuális rendszer részletfelismerő képessége számottevően rosszabb, mint fotopos körülmények között. A Purkinje jelenséget ismerve nyilvánvaló volt, hogy a fénysűrűség csökkenésével a V(λ) görbe maximumhelye fokozatosan eltolódik a rövidebb hullámhosszak felé, és némileg az alakja is módosul.
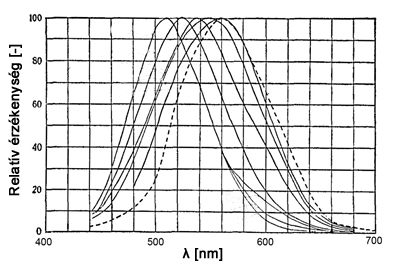
Ha a szkotopos láthatósági görbéből indulunk ki, azt tapasztalhatjuk, hogy a fénysűrűség növekedésével előbb a hosszabb hullámhosszak felé eső görbeszakaszon növekszik az érzékenység, majd a görbe maximumhelye ezután kezd csak a vörös tartomány felé vándorolni. A 2.26. ábra Walters és Wright által 1943-ban különböző fénysűrűség szintek mellett végzett világosság érzékelésre vonatkozó mérések eredményeként kapott görbéket mutatja.
Az már a mezopos látással kapcsolatos kísérletek kezdeti szakaszában nyilvánvalóvá vált, hogy az átmenet nem modellezhető a fotopos és szkotopos görbék egyszerű szuperpozíciójaként, más mechanizmusok is szerepet játszanak a köztes görbék lefutásának alakításában. Arra is hamar fény derült, hogy a vizsgálatok során alkalmazott látómező mérete értelemszerűen befolyásolja a kapott görbék lefolyását, ám a fénysűrűség csökkenésével némileg még foveális területen is megnő a rövidebb hullámhosszakra vett érzékenység, holott ezen a retinatartományon nem találhatóak pálcikák.
A mezopos világosság érzékelés mérésére alkalmas módszer meghatározása sem egyértelmű. Mivel a pálcikák és csapok együttes működésével a magnocelluláris és parvocelluláris idegpályák is szerepet játszanak a végső észlelet kialakításában, a villogásos fotometriával – ahol a parvocelluláris pályákon közlekedő jelek hatása nem érvényesül - és a közvetlen összehasonlításos módszerrel elvégzett mérések eredményei egymástól eltérnek, hiszen a közvetlen összehasonlításos módszer alkalmazása esetén heterokromatikus színegyeztetéskor fellép a korábban már említett Helmholtz-Kohlrausch effektus.
Ezen effektusok hatásainak jobb megértésével nyilvánvalóvá vált, hogy a mezopos átmenet nem lineáris folyamat. 1964-ben a CIE ezen nemlinearitások kezelésére és leírására vezette be az egyenértékű fénysűrűség fogalmát. Az egyenértékű fénysűrűség eredeti definíciója szerint „tetszőleges spektrális összetételű vizsgált mező egyenértékű fénysűrűségén azon összehasonlító mező fénysűrűségét értjük, melynek színhőmérséklete 2042 K, és világossága azonos a vizsgált mező világosságával”. A kandela definíciójának módosításakor az egyenértékű fénysűrűség definíciója is módosításra került, a 2042 K-es színhőmérsékletű összehasonlító fényforrás helyett az 540⋅1012 Hz-es monokromatikus sugárzással megvilágított mező került a definícióba.
Mindezeken túl, ugyanezen az 1964-es ülésen több, Walters és Wright görbéihez hasonló, adott fénysűrűség értékek mellett mért láthatósági függvény is elfogadásra került. A tématerület kutatása azonban inkább az egyenértékű fénysűrűség számításának metódusai felé mozdult el. Előbb Palmer dolgozott ki egy modellt, amely a fotopos és szkotopos fénysűrűségekből származtatja a mezopos egyenértékű fénysűrűség formulát, majd Kokoschka javasolt egy olyan számítási módot, amely mindhárom típusú csap receptor és a pálcikák jeleinek figyelembevételével definiálja a mezopos egyenértékű fénysűrűség értékét. Mindkét modelltípus zsákutcának bizonyult, hiszen a legújabb kutatások eredményeként előtérbe került fényérzékeny ganglion sejtek hatásait nélkülözik. Habár az napjainkban sem bizonyított, hogy létezik-e közvetlen kapcsolat ezen sejtek és a látókéreg között, a pupillareflex kialakításában és működésében bizonyosan részt vesznek, így befolyással bírnak a világosságérzékelés folyamatára.
A mezopos világosságérzékelés jelenlegi legmodernebb formuláját a CIE 2011-ben kiadott műszaki jelentése tartalmazza, amely a világosságmetrika kiegészítő rendszereként mezopos esetben is használható. A formula a korábban már ismertetett egyenértékű fénysűrűség fogalmát használja, és 2°-os foveális, vagy 10°-os perifériális látómezőre alkalmazható. A metrika figyelembe veszi a szkotopos és fotopos látásmechanizmusok sajátosságait, tekintettel van a kromatikus és akromatikus adaptációs folyamatok, valamint a parvocelluláris idegpályákon továbbított színi csatornák jeleinek hatásait is, melyhez mindhárom csap receptor által közvetített jel hozzájárul.
Mivel a kutatások szerint a pálcikák alkotta rendszer 10 cd/m2 fénysűrűség fölött telítésbe megy, a módszer itt a fotopos fénysűrűségnek megfelelő egyenértékű fénysűrűséget szolgáltatja. Az is ismert, hogy a csap receptorok csak 0,01 cd/m2 fölött kezdenek hozzájárulni a világosság érzékeléshez, így ezen érték alatt a formula által visszaadott eredmény a szkotopos fénysűrűséggel egyenértékű. A két fénysűrűség érték közötti tartományon az ismert mezopos átmeneti folyamatok hatásait is figyelembe vett, ennek megfelelően a (2.27. ábra) ábrán látható modell adja a legjobb lehetőséget a világosság érzékelés univerzális leírására.
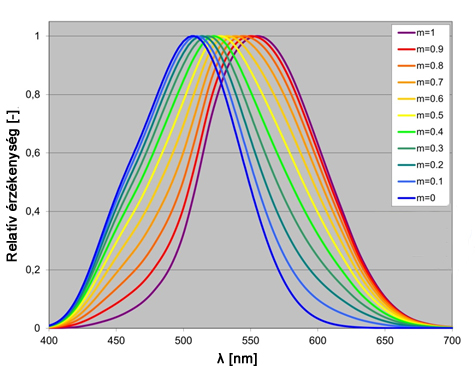
A mezopos világosság érzékelés metódusának pontosításán túl a kutatók figyelme az utóbbi években a vizuális teljesítmény fénysűrűség csökkenésével beálló változására irányult. A problémakört a modern fényforrások elterjedése tette különösen aktuálissá a közlekedéssel kapcsolatos vizuális feladatok kutatásában, hiszen a modern fémhalogén lámpák és LED-ek elterjedésével a korábban útvilágításban jellemzően alkalmazott nátriumlámpákénál jóval összetettebb színképi teljesítmény eloszlással rendelkező fényforrások kerültek piacra. A vizsgálatok tárgyát leginkább az képezte, hogy van-e eltérés a fent említett fényforrások alkalmazásával a mezopos körülmények között tapasztalható vizuális teljesítményben, ha a fotopos fénysűrűségek azonosak. A kutatások eredményeként a CIE a 2010-es vonatkozó műszaki jelentésében egy adaptív mezopos modellt tett közzé, amely a mezopos adaptációs fénysűrűséget és színképi érzékenységet a fotopos és szkotopos görbék összegéből származtatja, azonban beiktat egy fénysűrűségfüggő arányossági tagot az alábbiak szerint:
|
|
(2.1) |
Ahol L e az adaptációs sugársűrűség, V mes (λ) pedig:
|
|
(2.2) |
Ahol M(m) normalizáló függvény, amely V mes (λ) maximum értékét 1-re normálja, 0 ≤ m ≤ 1 adaptációs viszonyoktól függő arányossági tényező. Az m érték meghatározása az alábbiak szerint történik: L mes ≥ 5 cd/m2 esetén m = 1, L mes ≤ 0.005 cd/m2 esetén m = 0. A köztes tartományon m meghatározása iterációval történik, a következő módon:
|
|
(2.3) |
Ahol,
|
|
(2.4) |
Továbbá 0 ≤ m n ≤ 1, L p a fotopos, L s a szkotopos fénysűrűség, V’(λ 0 ) = 683/1700 λ 0 = 555 nm-es hullámhosszra, n az iterációs lépésszám, kiindulásként m 0 = 0.5 választandó, a = 0.767, b = 0.3334.
A fenti formulával kapott V mes (λ) függvényeket különböző m értékekre a 2.27. ábra mutatja.
Amennyiben a L s /L p fénysűrűség arány, amely megegyezik Φ s /Φ p szkotopos és fotopos fényáramok arányával, ismert, L mes mezopos fénysűrűség számítható. A szkotopos és fotopos viszonyok közötti fotometrikus arányszámot a fényforrásgyártók katalógusaikban jellemzően S/P értékként tüntetik fel.
Fontos megjegyezni, hogy habár a fentiekben szó esik fotopos, szkotopos és mezopos fotometriai mennyiségekről, ezek csak a mennyiségek megnevezésére vonatkozó előtagok, maguknak a mennyiségeknek és hozzájuk kapcsolódó mértékegységeknek nincsenek külön fotopos, szkotopos és mezopos megfelelői, azaz nincsen például mezopos cd/m2, vagy lux. Mezopos esetre a mennyiségek közül is kiemelendő a mezopos fényáram kifejezés, amelynek mértéke nem csak a fényforrás tulajdonságaitól, hanem az adaptációs állapottól is függ, így használata nehézkes, és nem is javasolt.
A formula használatát nehezíti, hogy egy adott útvilágítási vagy egyéb vizuális feladat esetén L mes értéke a látómező különböző pontjaiban nem állandó, helyről-helyre változik. Alkalmazását tovább bonyolítja, hogy jelenleg is kutatások tárgyát képezi, hogy hogyan határozzuk meg az adott feladathoz tartozó adaptációs állapotot, illetve hogy a különböző vizuális feladatok esetén a néző figyelme hova irányul, a nézési irány megváltozása hogyan van hatással az adaptációs állapot alakulására, valamint milyen befolyással van a körülményekre, ha az észlelő mozgásban van.
Összességében elmondható tehát, hogy a mezopos fotometria számos aspektusból komoly fontossággal bír a közlekedésbiztonság területén. Mivel a kérdéskör fiziológiai háttere rendkívül összetett, a látórendszer számos különböző működési metódusának együttese által alkotott mechanizmus, leírása és méréstechnikája is bonyolult összefüggésekre, mérési és számítási módszerekre vezet. Járműoptikában különösen fontos a mezopos tartományban vett vizuális teljesítmény alakulása a fénysűrűség változásával, hiszen ez számos éjszakai vezetéshez kapcsolódó kérdéskört, így műszaki megoldást és szabványosítási folyamatot is meghatároz vagy meg fog határozni a közeljövőben.
2.3.3.2. Káprázás és vakítás
Látórendszerünk dinamikatartománya rendkívül nagy átfogású, azonban határai így is elérhetőek, számos különböző általános vizuális körülmény között is, zavarva ezzel látásérzékelésük normális működését. Mivel éjszakai autóvezetéskor látórendszerünk nehezen definiálható állapotban működik, ekkor az ilyen zavaró hatásokra különösen érzékenyek vagyunk, így ezek részletesebb tárgyalása mindenképpen indokolt. A két jellegzetes vizuális zavaró hatás közül az egyikről alapjait tekintve már korábban is szó esett bizonyos látásmechanizmusok határátmeneteinek tárgyalásakor, ez a vakítás jelensége, amelyet a látótér nagy fénysűrűségű pontjai által telítésbe vitt receptorok jelképzésének pontatlansága okoz, és jellemzően a látótér foveális részén lép fel. A másik jelenség a káprázás, amelynek fundamentumai a vakításhoz hasonlóak, azonban maga a jelenség kissé más hatásmechanizmusok mentén értelmezhető.
2.3.3.2.1. Káprázás fajtái, kapcsolatuk az autóvezetéssel
A káprázás jelensége akkor lép fel, amikor a szem receptorainak maximális érzékenységét meghaladó, körülbelül 105 cd/m2 külső behatás, fénysűrűség éri a szemet, jellemzően szórt fény formájában vagy parafoveális területen, azaz látóterünk perifériális részén.
Az MSZ EN 12665 a káprázást a következő formában határozza meg:
„A látás kényelmetlensége és/vagy a tárgyak felismerhetőségének a csökkenése, a fénysűrűség szokatlan eloszlásának vagy szokatlan értékének, illetve a térben vagy időben fellépő igen erős kontrasztnak a következtében.”
Ez a szabvány azonban nem tesz különbséget a rontó és zavaró káprázás között. A világítástechnikában nemzetközileg a két jelenség között különbséget kell tenni.
A Nemzetközi Világítástechnikai Szótár alapján:
-
Rontó káprázás: Olyan káprázás, amely rontja a tárgyak látását anélkül, hogy szükségképpen kényelmetlenséget okozna. A szemgolyóban létrejövő fényszórásra vezethető vissza [5.] .
-
Zavaró káprázás: A kápráztató hatások közül azt nevezzük zavaró káprázásnak (vagy pszichológiai káprázásnak), amely látási kényelmetlenséget okoz anélkül, hogy szükségképpen rontaná a tárgy látását. Ilyen zavart okozhat, ha pl. egy nagy fénysűrűségű tárgy van a perifériás látás területén.
A zavaró káprázás objektív okait mind a mai napig nem sikerült felderíteni, annak vizsgálatát rendszerint csak kérdőíves kiértékelés alapján végzik, újabban mérési eljárásokat próbálnak az észleletekhez hozzárendelni, az ilyenkor fellépő hunyorítást alapul véve [6.] .
Az ezzel kapcsolatos korábbi kutatások két fő csoportra oszthatók:
-
pszichometrikus vizsgálatok (pl. VCP Visual Comfort Probability = látási komfort valószínűsége);
-
a káprázás fotometriai leírása.
A CIE az UGR (Unified Glare Rating) egységes káprázás érzékelési skálát deklarálta általánosan használt módszernek a zavaró káprázás meghatározásához, amelynek értéke 10-tõl 28-ig terjedhet a legkisebb kápráztató hatástól a legnagyobbig.
|
|
(2.5) |
ahol:
|
L b |
a háttér fénysűrűsége |
|
|
Ω |
térszög, amely alatt a világítótestek látszanak |
|
|
p² |
pozíció index |
|
|
l |
a világítótestek fénysűrűsége |
Az UGR formula nem a megvilágítást, hanem a háttér fénysűrűséget tartalmazza. (A helyiségben létesült átlagos megvilágítás természetesen befolyásolja a háttér fénysűrűséget).
Egy gyakorlati példa az UGR használatára:
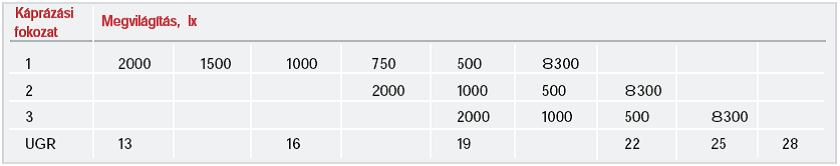
Kutatások [9.] , [10.] arra a következtetésre jutottak, hogy a zavaró káprázás-érzést elsősorban a kék-érzékeny (rövidhullámhossz-érzékeny) csapok közvetítik, de a fotopos színképi érzékenységnek is lényeges szerepe van.
A káprázást befolyásoló elsődleges tényezők:
-
a kápráztató fényforrás fénysűrűsége;
-
a térszög, amely alatt a szemlélési pontból nézve a kápráztató fényforrás látszik;
-
a kápráztató fényforrás elhelyezkedése a nézési irányhoz viszonyítva;
-
a kápráztató fényforrások száma, elrendezése;
-
a helyiség geometriai méretei;
-
valamint a környezet megvilágítása, ill. fénysűrűsége.
Közlekedésben gyakran előforduló káprázási jelenségek:
Egy izzólámpán, valamint egy Xe-lámpán végzett kísérlet során arra a következtetésre jutottak, hogy azonos zavaró káprázást beállítva, a fotopos vertikális megvilágítás (azaz a szem síkjában mérhető megvilágítás) mintegy másfélszer akkora volt halogén izzólámpa használatakor, mint Xe-lámpa esetén. Azaz, ha a Xe-lámpás és a halogén izzólámpás fényszóró esetén azonos fényeloszlást és útburkolat megvilágítást hoztak létre, akkor a Xe-lámpa által keltett zavaró káprázás nagyobb volt. Továbbá vizsgálták azt is [8.] , hogy ez a jelenség lehet-e az életkor illetve a fényforrások színének függvénye, s azt találták, hogy az idősebb és fiatalabb gépkocsivezetők különbözőképpen reagálnak a kétféle fényforrásra (2.28. ábra). Látható, hogy életkortól függetlenül a kékesebb árnyalatú gázkisülő lámpa erősebben kápráztat, de idősebb korban ez a hatás nagyobb.
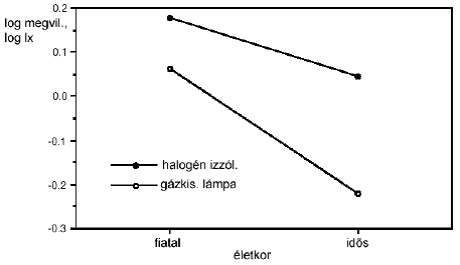
Az úttest kivilágítása annál kedvezőbb, minél nagyobb az átlagos megvilágítás erőssége, és minél kisebb az egyes helyek megvilágításában fennálló különbség. Ha az úttest egyes részeinek megvilágításában nagyok az eltérések, akkor a jó adaptációs képességű szem sem tudja követni a gyors fényerősség változásokat. Ezt a hatást mindenki ismeri, aki világos nappali közlekedés során szakszerűtlenül kivilágított alagúton hajtott át (2.29. ábra).

2.3.3.2.2. Vakítás jelensége, kapcsolata az autóvezetéssel
A vakítás a káprázáshoz hasonló optikai jelenség, de még fokozottabban veszélyes. Ilyen helyzet alakulhat ki a szembejövő jármű tompítatlan fényszóróival való találkozáskor. De olyan vakítás is felléphet, amelyet a síkfelületekről visszavert polarizált fény okoz (például vízfelszínről, autószélvédőről visszaverődő napfény).
A szem nappali receptorait, a csapokat ilyenkor maximális érzékenységüknél jóval nagyobb fénysűrűségű behatás éri, és ilyenkor a fotopos fotometria szabályai érvényüket vesztik. A vakítás
-
csökkenti a látáskomfortot
-
zavarja a színérzékelést
-
rontja a tér- és mélységérzékelést
-
extrém diszkomfortot okoz.
Vakítás esetében beszélhetünk fiziológiai és pszichológiai vakításról.
Hosszú másodpercekig tarthat, amíg az erős fénytől átmenetileg elvakított gépkocsivezető visszanyeri korábbi látóképességét. Amíg ez be nem következik, vakon és csak ösztönösen vezet, a baleseti veszély pedig óriási mértékben megnő. Az elvakított személy tulajdonképpen néhány másodpercig alkalmatlanná válik az autóvezetésre. A vakítási tilalmat viszonylag sokan megszegik, részint udvariatlanságból, ill. erőszakosságból, részint pedig a fényszórók hibás beállítása folytán.
A vakító jelenség a szabadban bárhol jelentkezhet:
-
vízfelszínen, havon, jégen
-
ködben, párában, szmogban
-
autók szélvédőjén, úttesten,ablakokon, tükröző felületeken.
A vakítás elleni védekezés egyedüli eszköze lehet a polarizált szemüvegek használata. A lencsék anyagára felvitt filmréteg sajátos tulajdonságainak köszönhetően a vízszintes felületekről visszaverődő fénysugarakat blokkolja, így védi a szemet a zavaró vakítástól (2.30. ábra).
